Zobacz slidy












Treść prezentacji
Slide 1
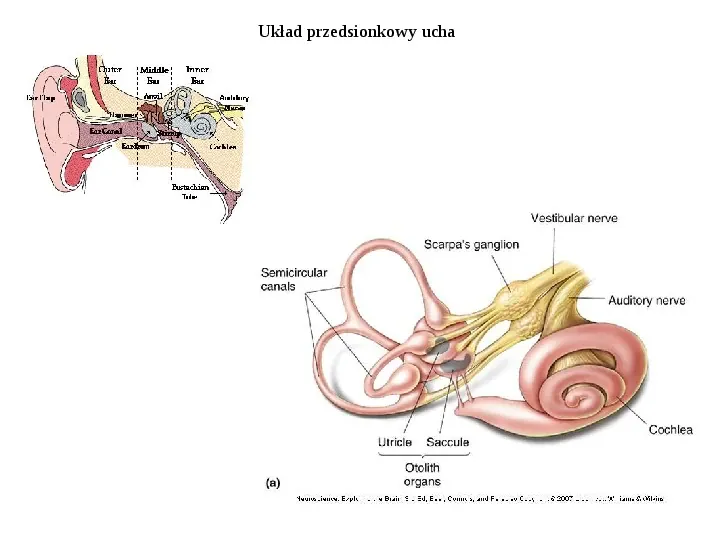
Układ przedsionkowy ucha
Slide 2
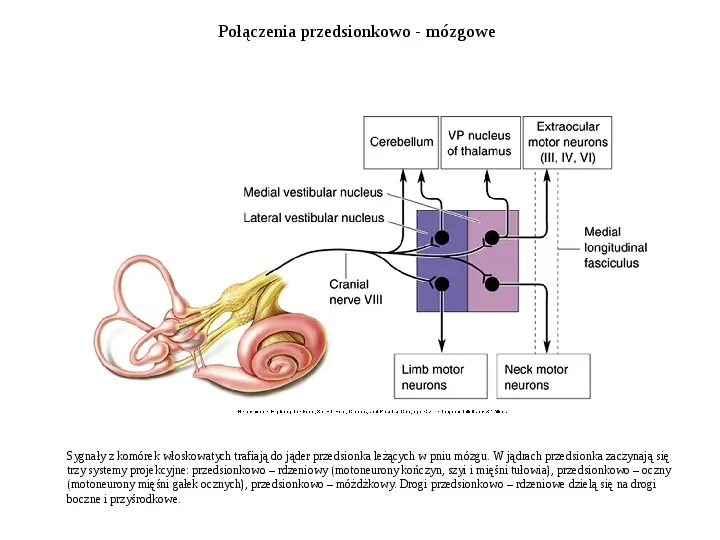
Połączenia przedsionkowo - mózgowe Sygnały z komórek włoskowatych trafiają do jąder przedsionka leżących w pniu mózgu. W jądrach przedsionka zaczynają się trzy systemy projekcyjne: przedsionkowo rdzeniowy (motoneurony kończyn, szyi i mięśni tułowia), przedsionkowo oczny (motoneurony mięśni gałek ocznych), przedsionkowo móżdżkowy. Drogi przedsionkowo rdzeniowe dzielą się na drogi boczne i przyśrodkowe.
Slide 3
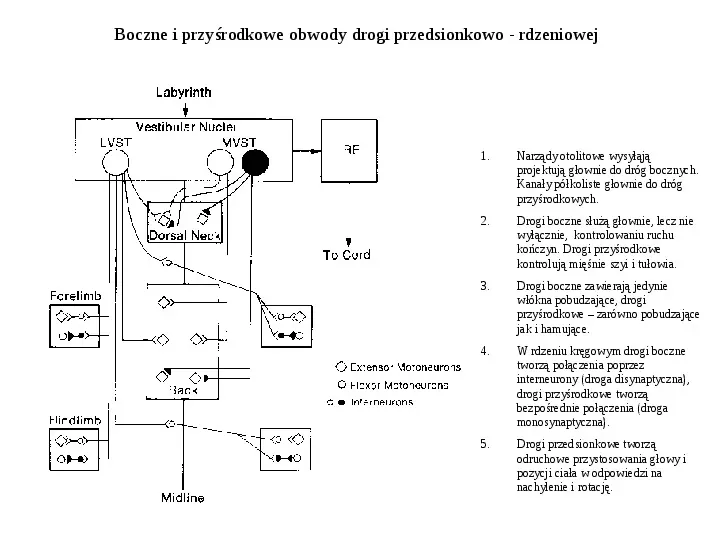
Boczne i przyśrodkowe obwody drogi przedsionkowo - rdzeniowej 1. Narządy otolitowe wysyłąją projektują głownie do dróg bocznych. Kanały półkoliste głownie do dróg przyśrodkowych. 2. Drogi boczne służą głownie, lecz nie wyłącznie, kontrolowaniu ruchu kończyn. Drogi przyśrodkowe kontrolują mięśnie szyi i tułowia. 3. Drogi boczne zawierają jedynie włókna pobudzające, drogi przyśrodkowe zarówno pobudzające jak i hamujące. 4. W rdzeniu kręgowym drogi boczne tworzą połączenia poprzez interneurony (droga disynaptyczna), drogi przyśrodkowe tworzą bezpośrednie połączenia (droga monosynaptyczna). 5. Drogi przedsionkowe tworzą odruchowe przystosowania głowy i pozycji ciała w odpowiedzi na nachylenie i rotację.
Slide 4
Współpraca odruchów szyjnych i błędnikowych w odruchowym przystosowaniu kończyn w odpowiedzi na nachylenie ciała. Sygnały przedsionkowe (błędnikowe) określają położenie głowy, które zmienia się w wyniku nachylenia i obrotu głowy i ciała. Odruchy te działają na mięśnie szyi by przywrócić głowę do pozycji pozycji normalnej i na mięśnie kończyn by utrzymać równowagę Naciąg mięsni szyjnych również wytwarza odruchowe skurcze mięśni szyjnych i kończyn. Odruchy przedsionkowe i szyjne mogą działać synergistycznie lub antagonistycznie
Slide 5
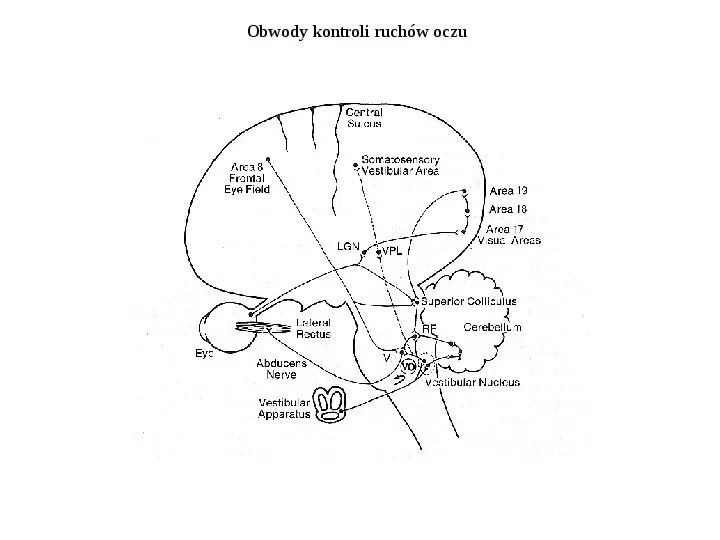
Obwody kontroli ruchów oczu
Slide 6
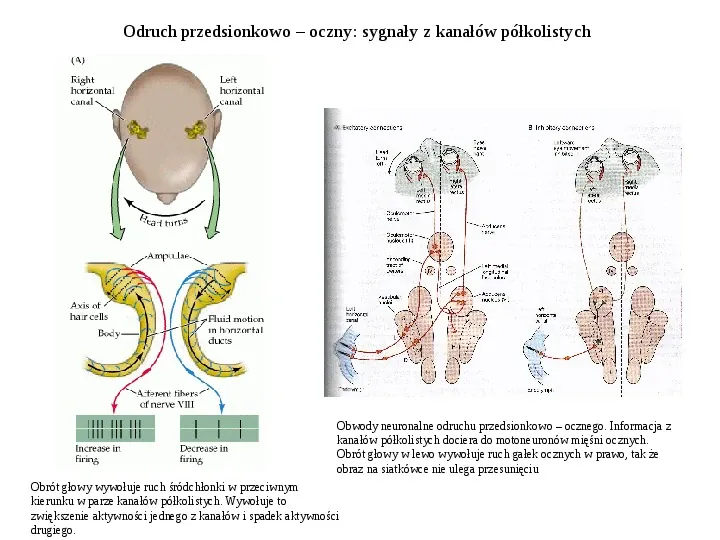
Odruch przedsionkowo oczny: sygnały z kanałów półkolistych Obwody neuronalne odruchu przedsionkowo ocznego. Informacja z kanałów półkolistych dociera do motoneuronów mięśni ocznych. Obrót głowy w lewo wywołuje ruch gałek ocznych w prawo, tak że obraz na siatkówce nie ulega przesunięciu Obrót głowy wywołuje ruch śródchłonki w przeciwnym kierunku w parze kanałów półkolistych. Wywołuje to zwiększenie aktywności jednego z kanałów i spadek aktywności drugiego.
Slide 7
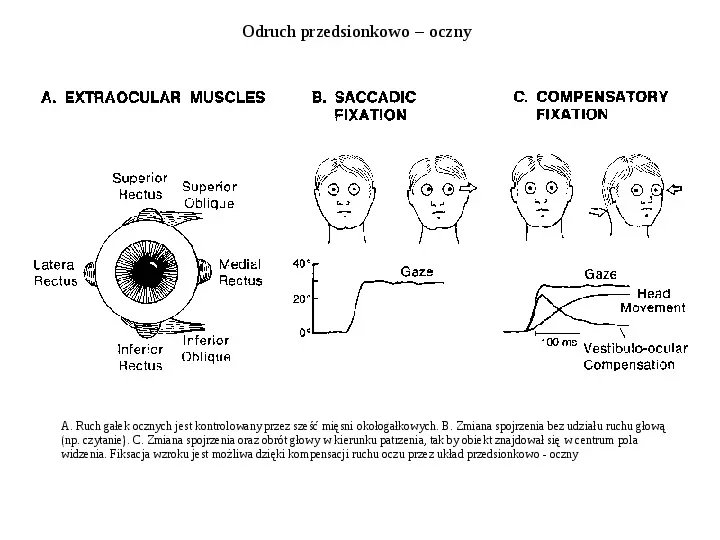
Odruch przedsionkowo oczny A. Ruch gałek ocznych jest kontrolowany przez sześć mięsni okołogałkowych. B. Zmiana spojrzenia bez udziału ruchu głową (np. czytanie). C. Zmiana spojrzenia oraz obrót głowy w kierunku patrzenia, tak by obiekt znajdował się w centrum pola widzenia. Fiksacja wzroku jest możliwa dzięki kompensacji ruchu oczu przez układ przedsionkowo - oczny
Slide 8
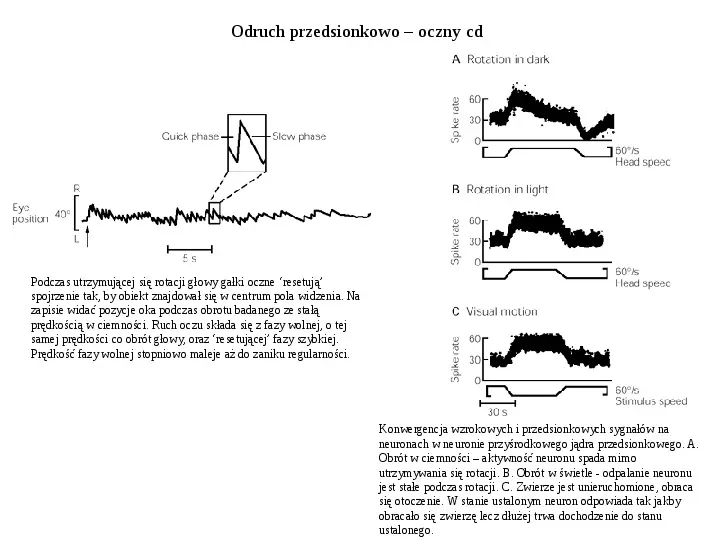
Odruch przedsionkowo oczny cd Podczas utrzymującej się rotacji głowy gałki oczne resetują spojrzenie tak, by obiekt znajdował się w centrum pola widzenia. Na zapisie widać pozycje oka podczas obrotu badanego ze stałą prędkością w ciemności. Ruch oczu składa się z fazy wolnej, o tej samej prędkości co obrót głowy, oraz resetującej fazy szybkiej. Prędkość fazy wolnej stopniowo maleje aż do zaniku regularności. Konwergencja wzrokowych i przedsionkowych sygnałów na neuronach w neuronie przyśrodkowego jądra przedsionkowego. A. Obrót w ciemności aktywność neuronu spada mimo utrzymywania się rotacji. B. Obrót w świetle - odpalanie neuronu jest stałe podczas rotacji. C. Zwierze jest unieruchomione, obraca się otoczenie. W stanie ustalonym neuron odpowiada tak jakby obracało się zwierzę lecz dłużej trwa dochodzenie do stanu ustalonego.
Slide 9
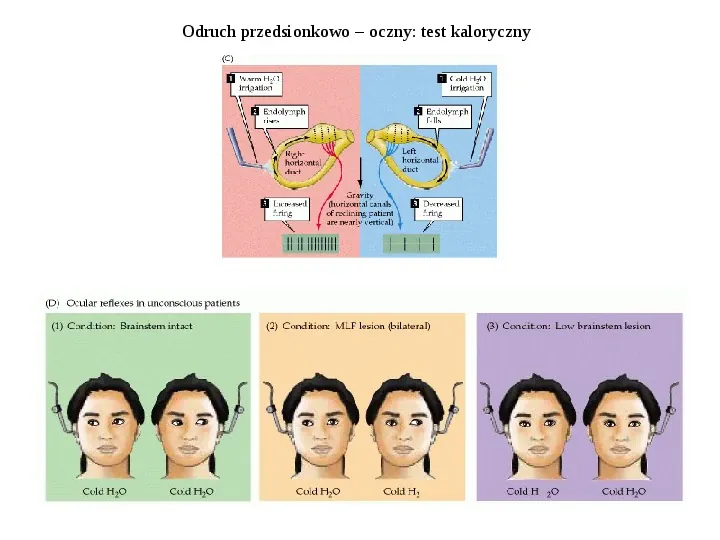
Odruch przedsionkowo oczny: test kaloryczny
Slide 10
Stan nieważkości 1. Wrażenie raczej przyjemne 2. Szybka adaptacja(!) 3. Procesy adaptacyjne widoczne po powrocie na Ziemie, zanikające z czasem.
Slide 11
Słuch
Slide 12
Dźwięki Zakres słyszalnych dźwięków u wybranych zwierząt.
Slide 13
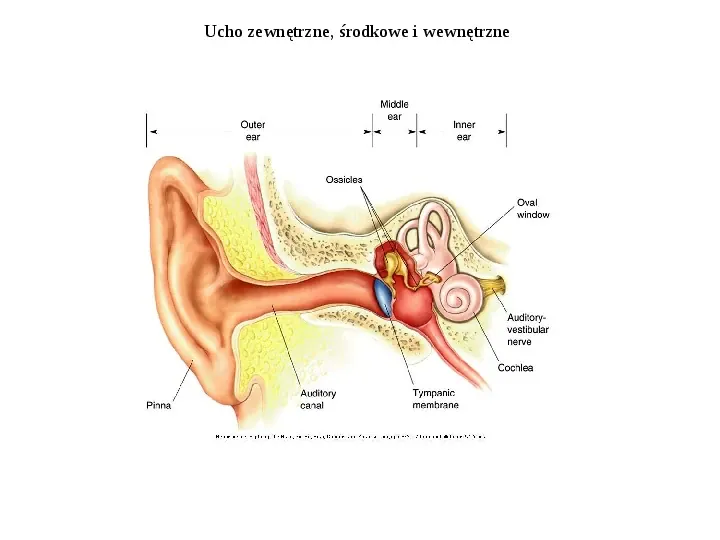
Ucho zewnętrzne, środkowe i wewnętrzne
Slide 14
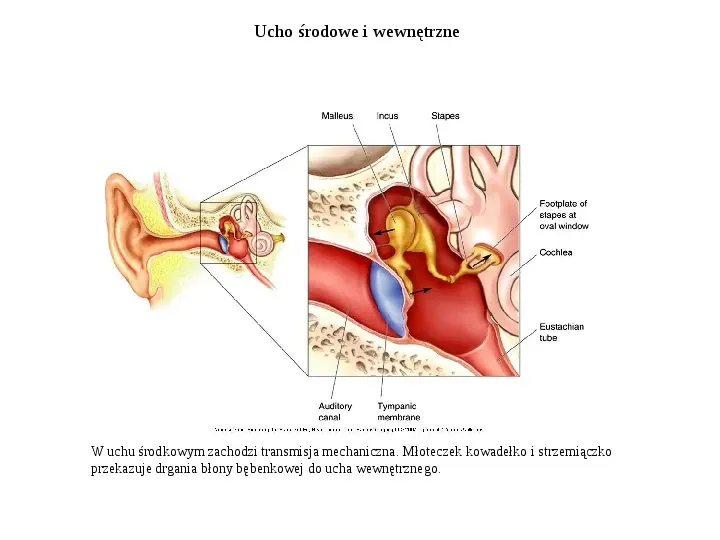
Ucho środowe i wewnętrzne W uchu środkowym zachodzi transmisja mechaniczna. Młoteczek kowadełko i strzemiączko przekazuje drgania błony bębenkowej do ucha wewnętrznego.
Slide 15
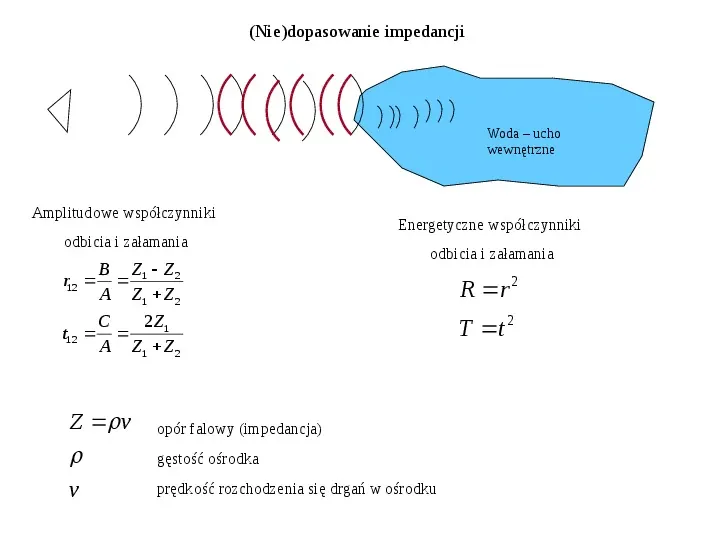
(Nie)dopasowanie impedancji Woda ucho wewnętrzne Amplitudowe współczynniki odbicia i załamania r12 B Z1 Z 2 A Z1 Z 2 t12 C 2Z1 A Z1 Z 2 Z v v Energetyczne współczynniki odbicia i załamania opór falowy (impedancja) gęstość ośrodka prędkość rozchodzenia się drgań w ośrodku R r 2 T t 2
Slide 16
(Nie)dopasowanie impedancji cd. powietrza 1.21 kgm3 wody 103 kgm3 v powietrza 340 ms vwody 1480 ms Energetyczne współczynniki odbicia i załamania R 0.9989 T 1 R 0.0011 Przechodzi 11000 energii fali: strata 30 dB
Slide 17
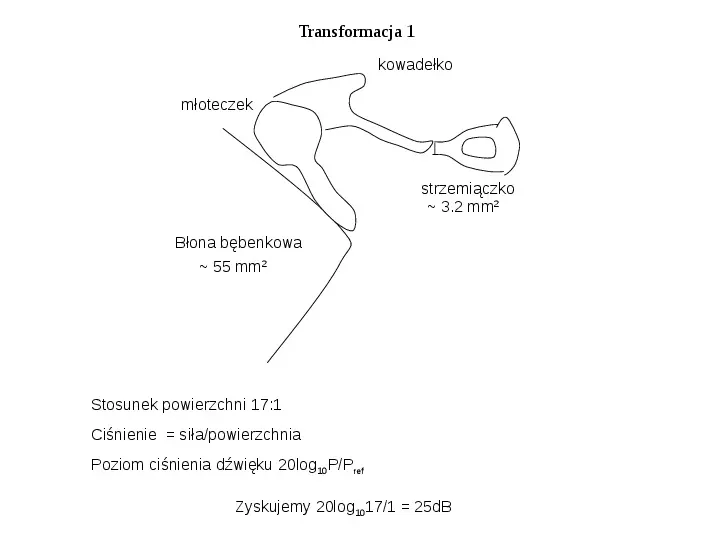
Transformacja 1 kowadełko młoteczek strzemiączko 3.2 mm2 Błona bębenkowa 55 mm2 Stosunek powierzchni 17:1 Ciśnienie siłapowierzchnia Poziom ciśnienia dźwięku 20log10PPref Zyskujemy 20log10171 25dB
Slide 18
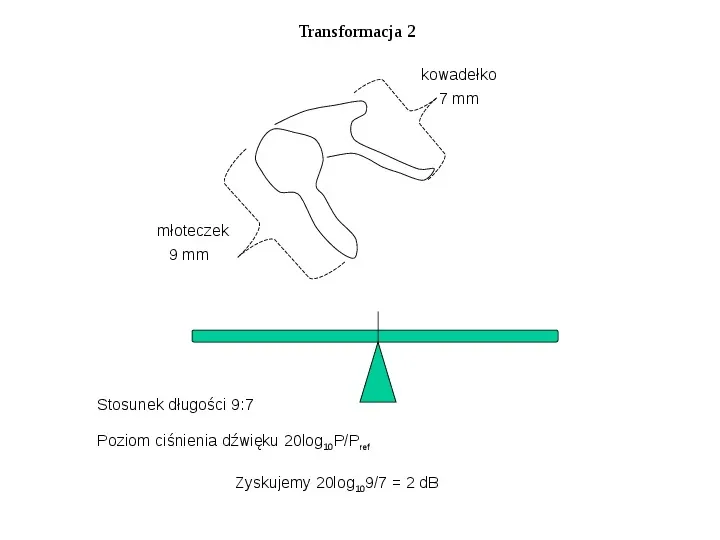
Transformacja 2 kowadełko 7 mm młoteczek 9 mm Stosunek długości 9:7 Poziom ciśnienia dźwięku 20log10PPref Zyskujemy 20log1097 2 dB
Slide 19
Amplituda drgań powietrza dla progu słyszalności ( x, t ) A cos(kx t ) Akustyczna fala biegnąca 2 1 2 2 2 E 2 xS xSA sin (kx t ) 2 t Energia fali (Epot Ekin) dE vdtSA2 2 1 2 I sin (kx t ) vA2 2 dtS dtS 2 Natężenie fali I I 0 10 12 Wm 2 Próg słyszalności 0 dB 1 A 2 Amplituda fali 2I v 1 2 10 12 A 1.26 10 11 m 2764 1.29 340 A średnica atomu ( 10-10 m) Amplituda fali dźwiękowej o częstości 440 Hz
Slide 20
Natężenie dźwięku i poziom natężenia dźwięku Ludzkie ucho odbiera natężenia z zakresu wartości 10-12 100 Wm2. Głośność (poziom natężenia) dźwięku jest zdefiniowana jako: (10dB) log gdzie: poziom natężenia dźwięku I natężenie dźwięku I0 wartość odniesienia, wynosząca 1012 Wm2 I I0 Czasami, mówi się też o poziomie ciśnienia dźwięku: (20dB)log p p0 Zalety skali decybelowej: 1. Łatwiejszy zapis dużego zakresu 2. Łatwiejsze obliczenia (wzmocnienie dodawanie, osłabienie odejmowanie). 3. Ogólnie przyjęta skala międzynarodowa 4. Wartość zero odpowiada ludzkiemu progowi słyszalności. 5. Zgodna z fizjologia zmysłu słuchu.
Slide 21
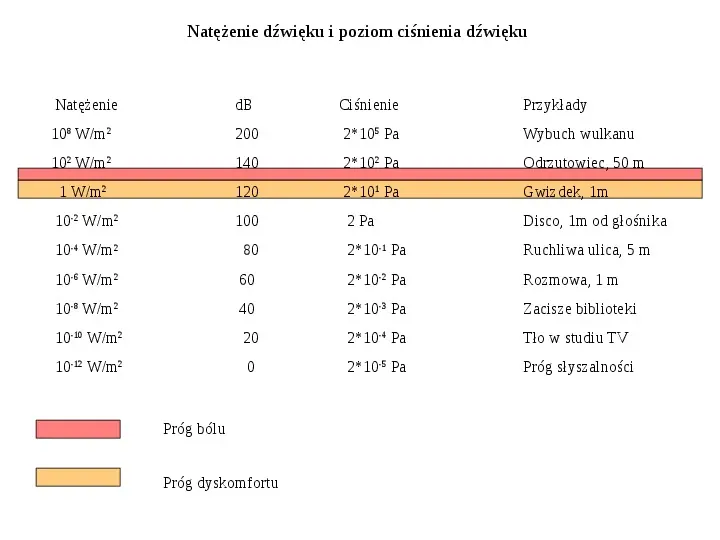
Natężenie dźwięku i poziom ciśnienia dźwięku Natężenie dB Ciśnienie Przykłady 108 Wm2 200 2105 Pa Wybuch wulkanu 102 Wm2 140 2102 Pa Odrzutowiec, 50 m 1 Wm2 120 2101 Pa Gwizdek, 1m 10-2 Wm2 100 2 Pa Disco, 1m od głośnika 10-4 Wm2 80 210-1 Pa Ruchliwa ulica, 5 m 10-6 Wm2 60 210-2 Pa Rozmowa, 1 m 10-8 Wm2 40 210-3 Pa Zacisze biblioteki 10-10 Wm2 20 210-4 Pa Tło w studiu TV 10-12 Wm2 0 210-5 Pa Próg słyszalności Próg bólu Próg dyskomfortu
Slide 22
Jak działa ucho?
Slide 23
Jak działa ucho?
Slide 24
Jak działa ucho?
Slide 25
Jak działa ucho?
Slide 26
Jak działa ucho?
Slide 27
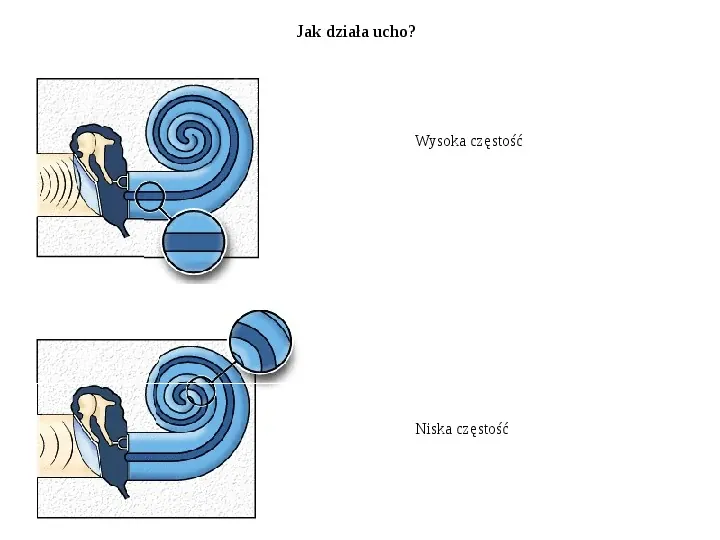
Jak działa ucho? Wysoka częstość Niska częstość
Slide 28
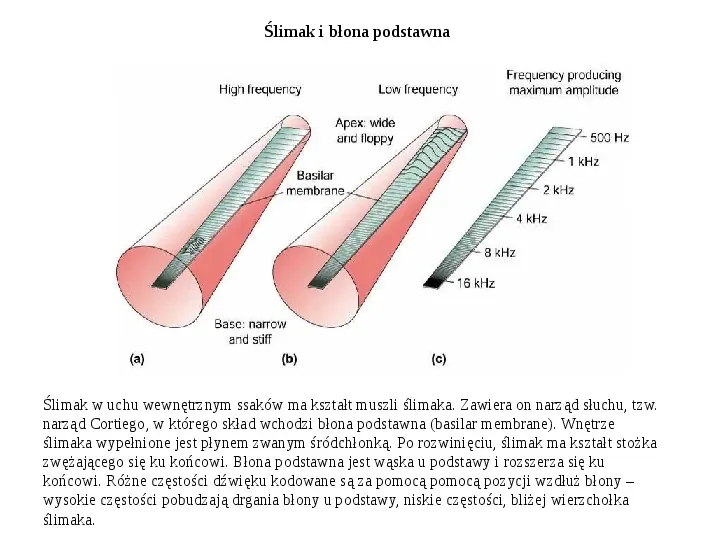
Ślimak i błona podstawna Ślimak w uchu wewnętrznym ssaków ma kształt muszli ślimaka. Zawiera on narząd słuchu, tzw. narząd Cortiego, w którego skład wchodzi błona podstawna (basilar membrane). Wnętrze ślimaka wypełnione jest płynem zwanym śródchłonką. Po rozwinięciu, ślimak ma kształt stożka zwężającego się ku końcowi. Błona podstawna jest wąska u podstawy i rozszerza się ku końcowi. Różne częstości dźwięku kodowane są za pomocą pomocą pozycji wzdłuż błony wysokie częstości pobudzają drgania błony u podstawy, niskie częstości, bliżej wierzchołka ślimaka.
Slide 29
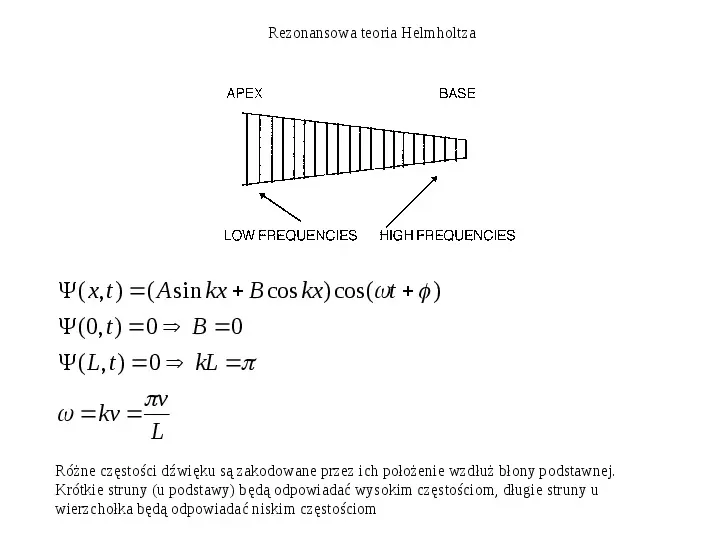
Rezonansowa teoria Helmholtza ( x, t ) ( A sin kx B cos kx) cos(t ) (0, t ) 0 B 0 ( L, t ) 0 kL v kv L Różne częstości dźwięku są zakodowane przez ich położenie wzdłuż błony podstawnej. Krótkie struny (u podstawy) będą odpowiadać wysokim częstościom, długie struny u wierzchołka będą odpowiadać niskim częstościom
Slide 30
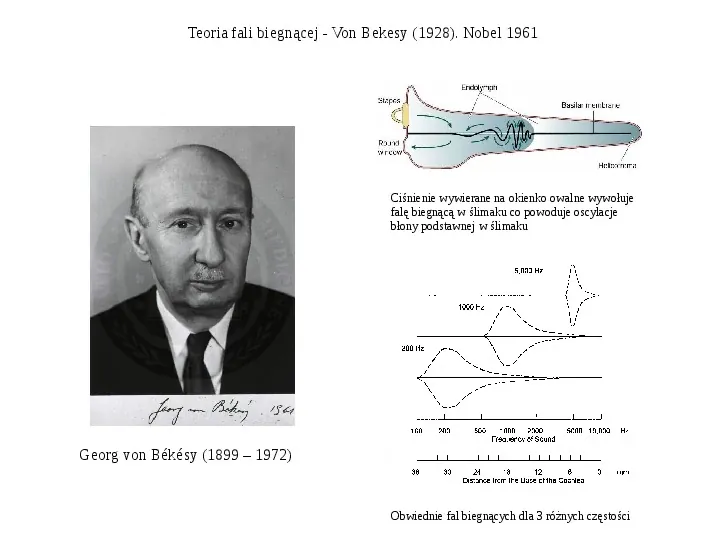
Teoria fali biegnącej - Von Bekesy (1928). Nobel 1961 Ciśnienie wywierane na okienko owalne wywołuje falę biegnącą w ślimaku co powoduje oscylacje błony podstawnej w ślimaku Georg von Békésy (1899 1972) Obwiednie fal biegnących dla 3 różnych częstości
Slide 31
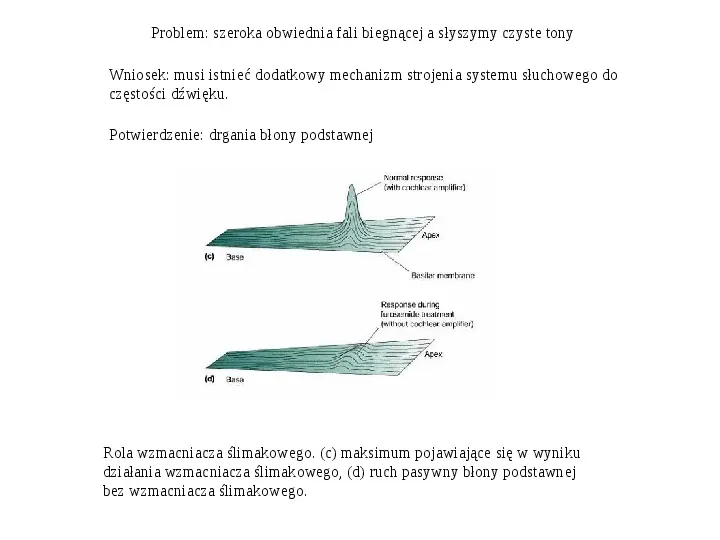
Problem: szeroka obwiednia fali biegnącej a słyszymy czyste tony Wniosek: musi istnieć dodatkowy mechanizm strojenia systemu słuchowego do częstości dźwięku. Potwierdzenie: drgania błony podstawnej Rola wzmacniacza ślimakowego. (c) maksimum pojawiające się w wyniku działania wzmacniacza ślimakowego, (d) ruch pasywny błony podstawnej bez wzmacniacza ślimakowego.
Slide 32
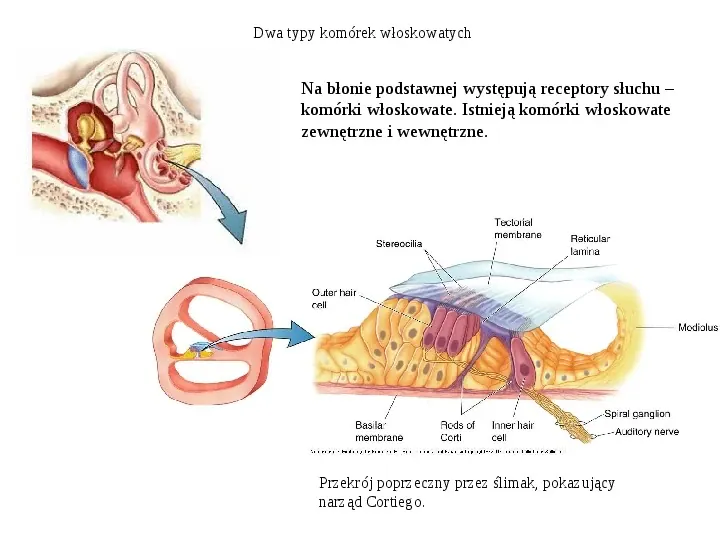
Dwa typy komórek włoskowatych Na błonie podstawnej występują receptory słuchu komórki włoskowate. Istnieją komórki włoskowate zewnętrzne i wewnętrzne. Przekrój poprzeczny przez ślimak, pokazujący narząd Cortiego.
Slide 33
Dwa typy komórek włoskowatych Mikrografie ze skaningowego mikroskopu elektronowego ukazujące ułożenie komórek włosowatych wewnętrznych i zewnętrznych oraz ich rzęsek. Komórki zewnętrzne ułożone są w trzy rzędy, a ich rzęski ułożone są w kształcie V. Komórki wewnętrzne leżą w jednym rzędzie.
Slide 34
Organizacja i własności wewnętrznych i zewnętrznych komórek włoskowatych
Slide 35
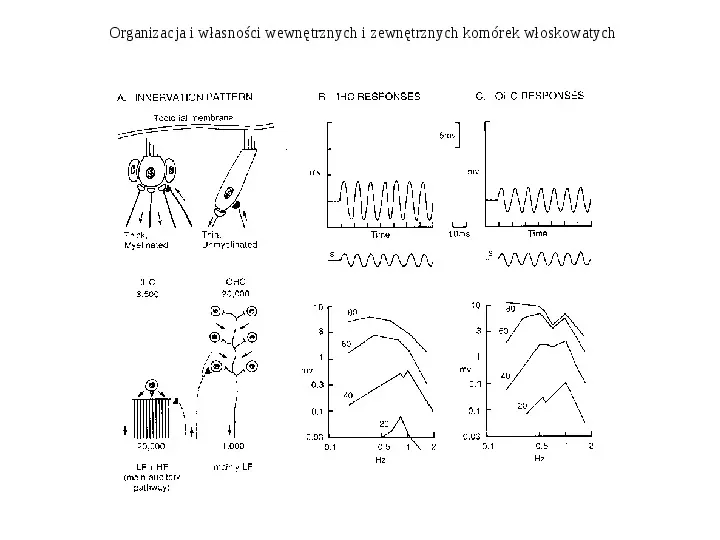
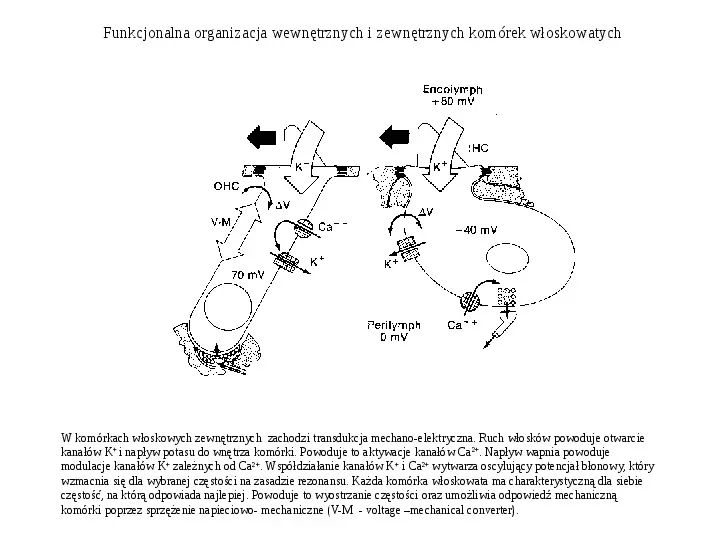
Funkcjonalna organizacja wewnętrznych i zewnętrznych komórek włoskowatych W komórkach włoskowych zewnętrznych zachodzi transdukcja mechano-elektryczna. Ruch włosków powoduje otwarcie kanałów K i napływ potasu do wnętrza komórki. Powoduje to aktywacje kanałów Ca2. Napływ wapnia powoduje modulacje kanałów K zależnych od Ca2. Współdziałanie kanałów K i Ca2 wytwarza oscylujący potencjał błonowy, który wzmacnia się dla wybranej częstości na zasadzie rezonansu. Każda komórka włoskowata ma charakterystyczną dla siebie częstość, na którą odpowiada najlepiej. Powoduje to wyostrzanie częstości oraz umożliwia odpowiedź mechaniczną komórki poprzez sprzężenie napieciowo- mechaniczne (V-M - voltage mechanical converter).
Slide 36
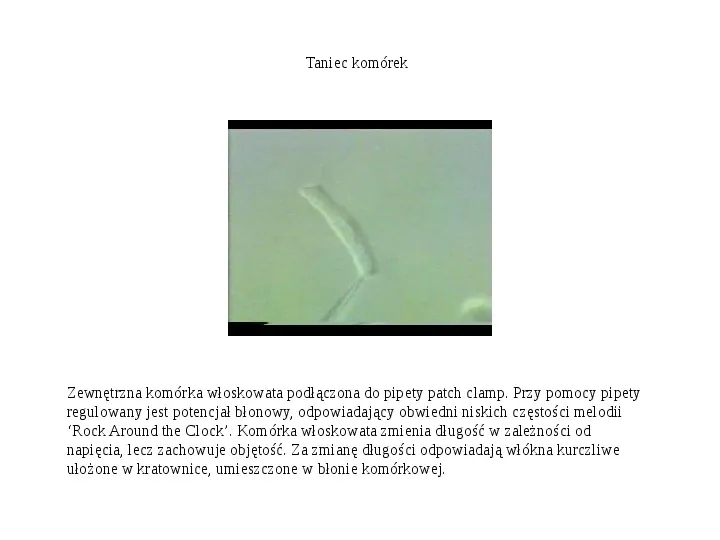
Taniec komórek Zewnętrzna komórka włoskowata podłączona do pipety patch clamp. Przy pomocy pipety regulowany jest potencjał błonowy, odpowiadający obwiedni niskich częstości melodii Rock Around the Clock. Komórka włoskowata zmienia długość w zależności od napięcia, lecz zachowuje objętość. Za zmianę długości odpowiadają włókna kurczliwe ułożone w kratownice, umieszczone w błonie komórkowej.
Slide 37
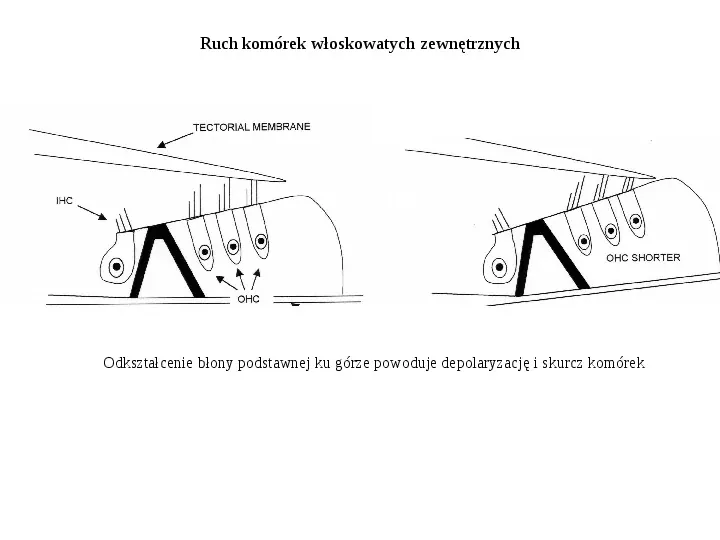
Ruch komórek włoskowatych zewnętrznych Odkształcenie błony podstawnej ku górze powoduje depolaryzację i skurcz komórek
Slide 38
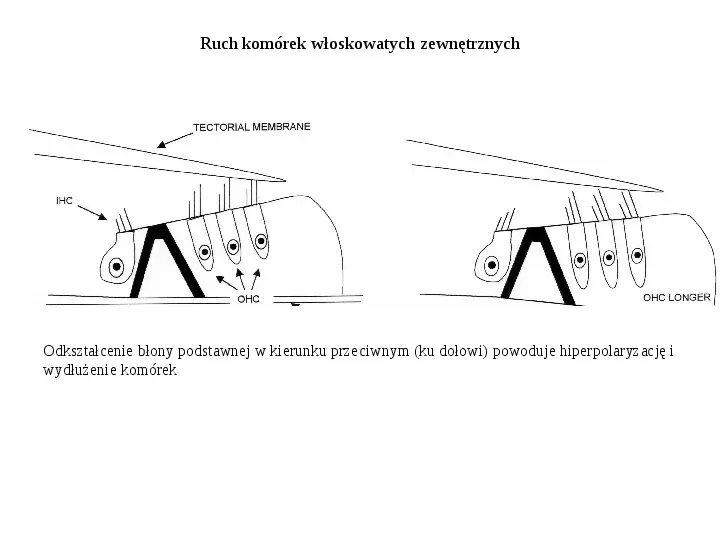
Ruch komórek włoskowatych zewnętrznych Odkształcenie błony podstawnej w kierunku przeciwnym (ku dołowi) powoduje hiperpolaryzację i wydłużenie komórek
Slide 39
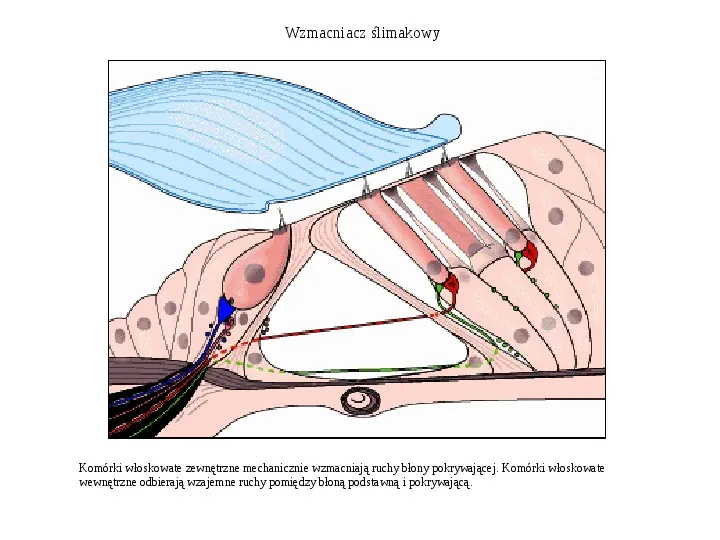
Wzmacniacz ślimakowy Komórki włoskowate zewnętrzne mechanicznie wzmacniają ruchy błony pokrywającej. Komórki włoskowate wewnętrzne odbierają wzajemne ruchy pomiędzy błoną podstawną i pokrywającą.
Slide 40
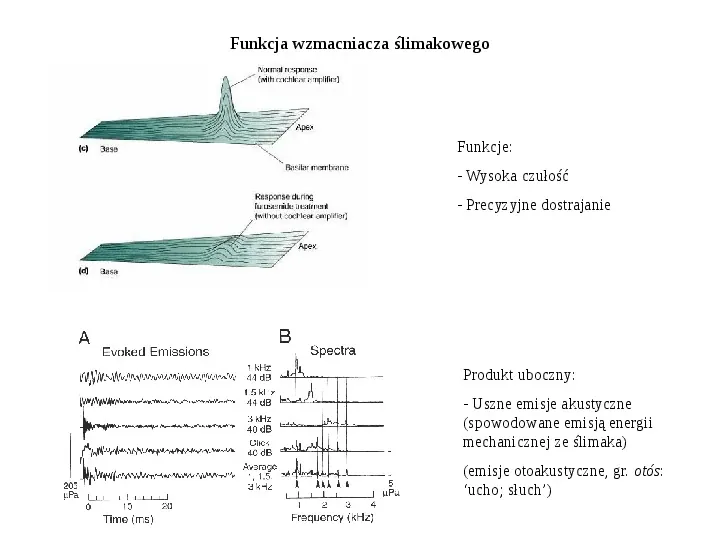
Funkcja wzmacniacza ślimakowego Funkcje: - Wysoka czułość - Precyzyjne dostrajanie Produkt uboczny: - Uszne emisje akustyczne (spowodowane emisją energii mechanicznej ze ślimaka) (emisje otoakustyczne, gr. otós: ucho; słuch)
Slide 41
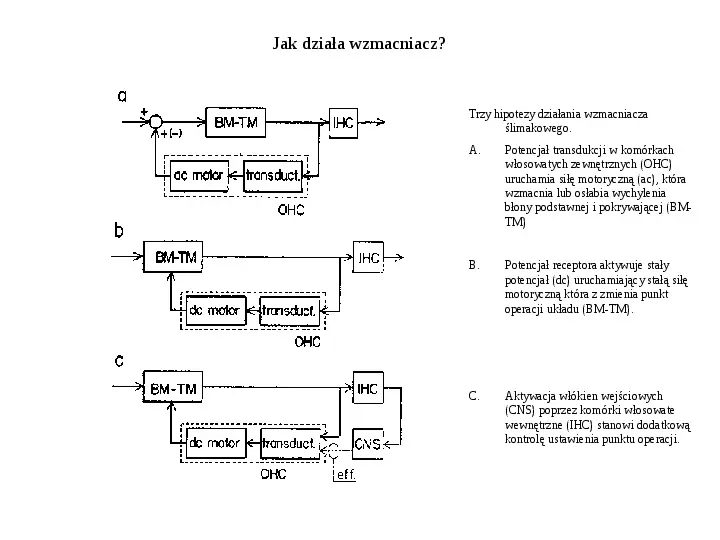
Jak działa wzmacniacz? Trzy hipotezy działania wzmacniacza ślimakowego. A. Potencjał transdukcji w komórkach włosowatych zewnętrznych (OHC) uruchamia siłę motoryczną (ac), która wzmacnia lub osłabia wychylenia błony podstawnej i pokrywającej (BMTM) B. Potencjał receptora aktywuje stały potencjał (dc) uruchamiający stałą siłę motoryczną która z zmienia punkt operacji układu (BM-TM). C. Aktywacja włókien wejściowych (CNS) poprzez komórki włosowate wewnętrzne (IHC) stanowi dodatkową kontrolę ustawienia punktu operacji.
Slide 42
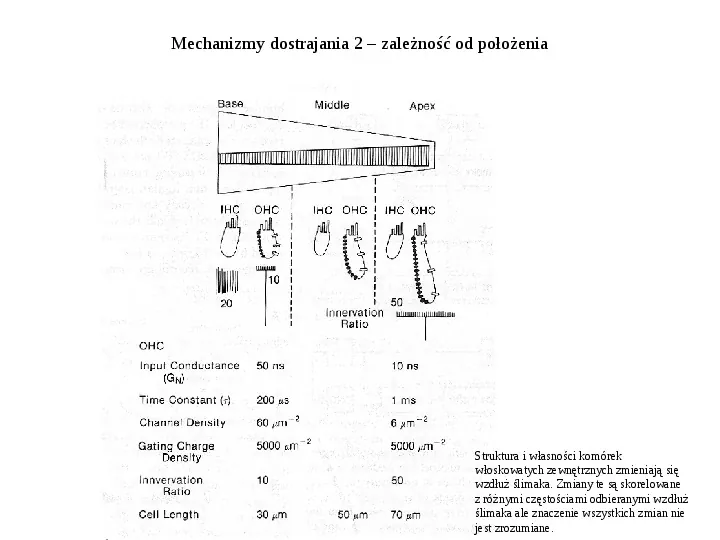
Mechanizmy dostrajania 2 zależność od położenia Struktura i własności komórek włoskowatych zewnętrznych zmieniają się wzdłuż ślimaka. Zmiany te są skorelowane z różnymi częstościami odbieranymi wzdłuż ślimaka ale znaczenie wszystkich zmian nie jest zrozumiane.
Slide 43
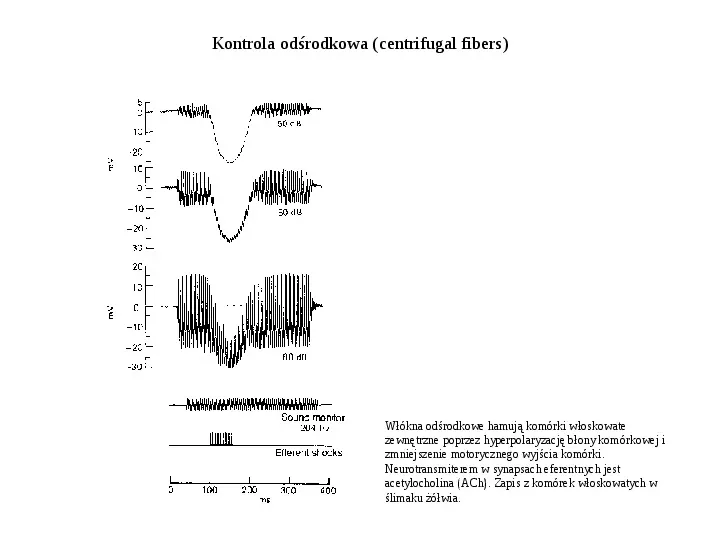
Kontrola odśrodkowa (centrifugal fibers) Włókna odśrodkowe hamują komórki włoskowate zewnętrzne poprzez hyperpolaryzację błony komórkowej i zmniejszenie motorycznego wyjścia komórki. Neurotransmiterem w synapsach eferentnych jest acetylocholina (ACh). Zapis z komórek włoskowatych w ślimaku żółwia.
Slide 44
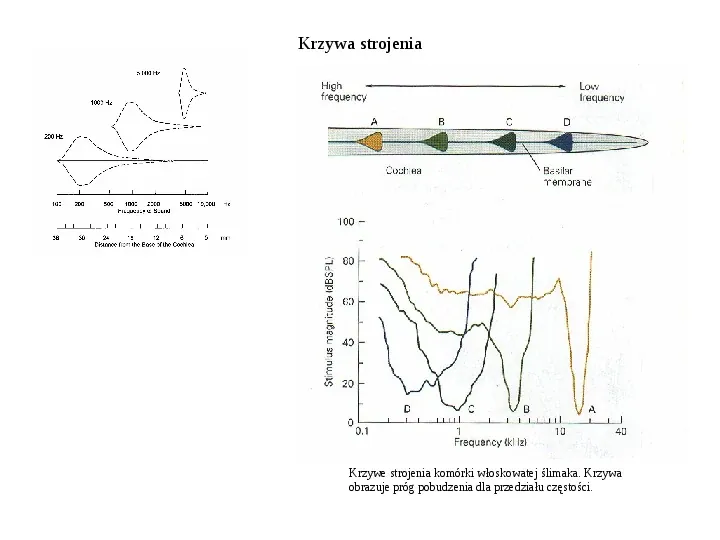
Krzywa strojenia Krzywe strojenia komórki włoskowatej ślimaka. Krzywa obrazuje próg pobudzenia dla przedziału częstości.
Slide 45
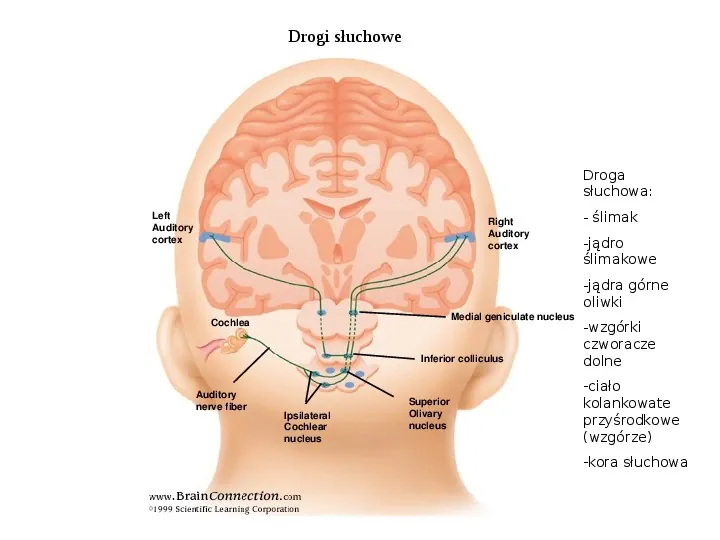
Drogi słuchowe Droga słuchowa: Left Auditory cortex Right Auditory cortex - ślimak -jądro ślimakowe -jądra górne oliwki Medial geniculate nucleus Cochlea Inferior colliculus Auditory nerve fiber Ipsilateral Cochlear nucleus Superior Olivary nucleus -wzgórki czworacze dolne -ciało kolankowate przyśrodkowe (wzgórze) -kora słuchowa
Slide 46
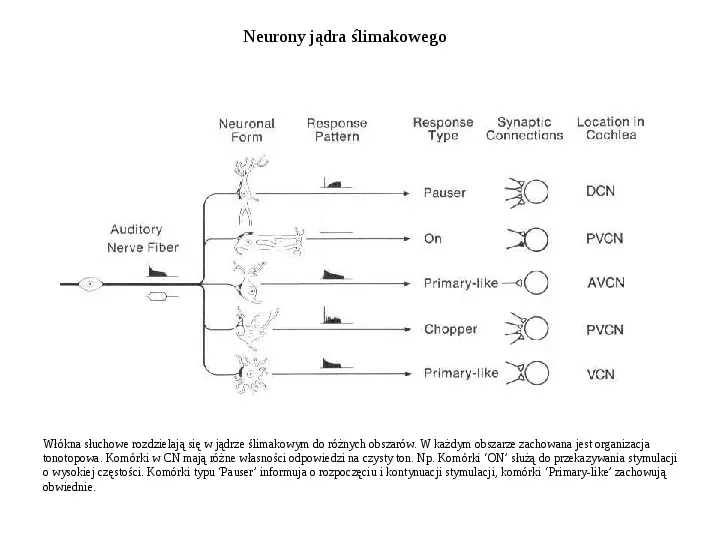
Neurony jądra ślimakowego Włókna słuchowe rozdzielają się w jądrze ślimakowym do różnych obszarów. W każdym obszarze zachowana jest organizacja tonotopowa. Komórki w CN mają różne własności odpowiedzi na czysty ton. Np. Komórki ON służą do przekazywania stymulacji o wysokiej częstości. Komórki typu Pauser informuja o rozpoczęciu i kontynuacji stymulacji, komórki Primary-like zachowują obwiednie.
Slide 47
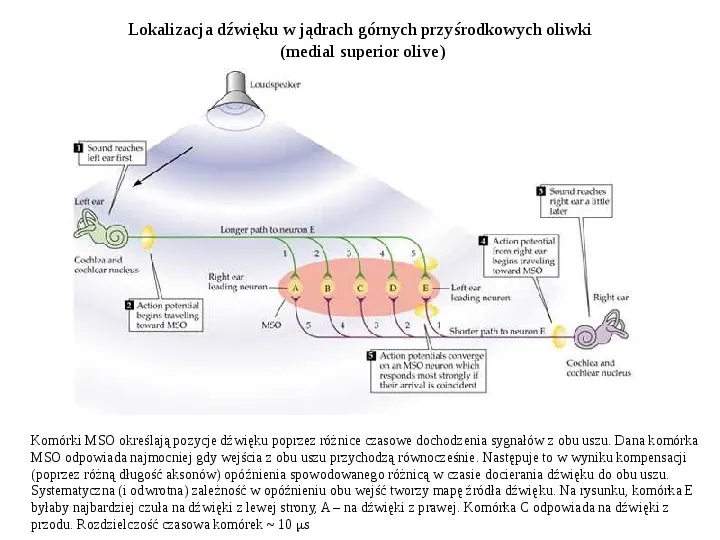
Lokalizacja dźwięku w jądrach górnych przyśrodkowych oliwki (medial superior olive) Komórki MSO określają pozycje dźwięku poprzez różnice czasowe dochodzenia sygnałów z obu uszu. Dana komórka MSO odpowiada najmocniej gdy wejścia z obu uszu przychodzą równocześnie. Następuje to w wyniku kompensacji (poprzez różną długość aksonów) opóźnienia spowodowanego różnicą w czasie docierania dźwięku do obu uszu. Systematyczna (i odwrotna) zależność w opóźnieniu obu wejść tworzy mapę źródła dźwięku. Na rysunku, komórka E byłaby najbardziej czuła na dźwięki z lewej strony, A na dźwięki z prawej. Komórka C odpowiada na dźwięki z przodu. Rozdzielczość czasowa komórek 10 s
Slide 48
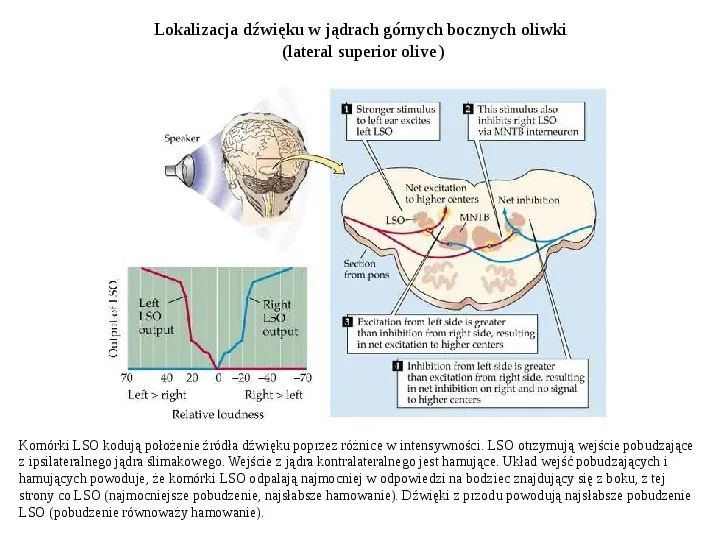
Lokalizacja dźwięku w jądrach górnych bocznych oliwki (lateral superior olive) Komórki LSO kodują położenie źródła dźwięku poprzez różnice w intensywności. LSO otrzymują wejście pobudzające z ipsilateralnego jądra ślimakowego. Wejście z jądra kontralateralnego jest hamujące. Układ wejść pobudzających i hamujących powoduje, że komórki LSO odpalają najmocniej w odpowiedzi na bodziec znajdujący się z boku, z tej strony co LSO (najmocniejsze pobudzenie, najsłabsze hamowanie). Dźwięki z przodu powodują najsłabsze pobudzenie LSO (pobudzenie równoważy hamowanie).
Slide 49
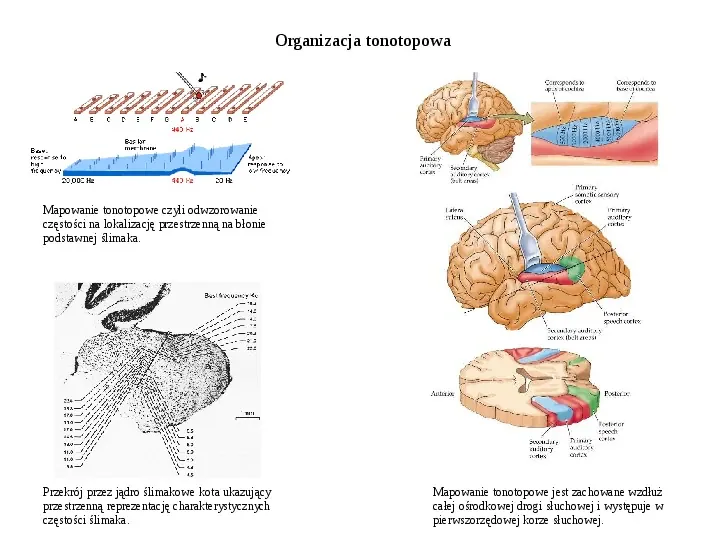
Organizacja tonotopowa Mapowanie tonotopowe czyli odwzorowanie częstości na lokalizację przestrzenną na błonie podstawnej ślimaka. Przekrój przez jądro ślimakowe kota ukazujący przestrzenną reprezentację charakterystycznych częstości ślimaka. Mapowanie tonotopowe jest zachowane wzdłuż całej ośrodkowej drogi słuchowej i występuje w pierwszorzędowej korze słuchowej.
Slide 50
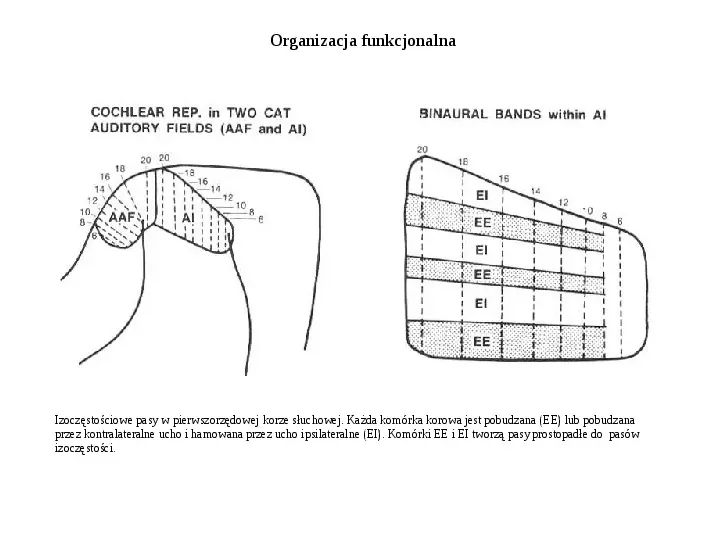
Organizacja funkcjonalna Izoczęstościowe pasy w pierwszorzędowej korze słuchowej. Każda komórka korowa jest pobudzana (EE) lub pobudzana przez kontralateralne ucho i hamowana przez ucho ipsilateralne (EI). Komórki EE i EI tworzą pasy prostopadłe do pasów izoczęstości.
Slide 51
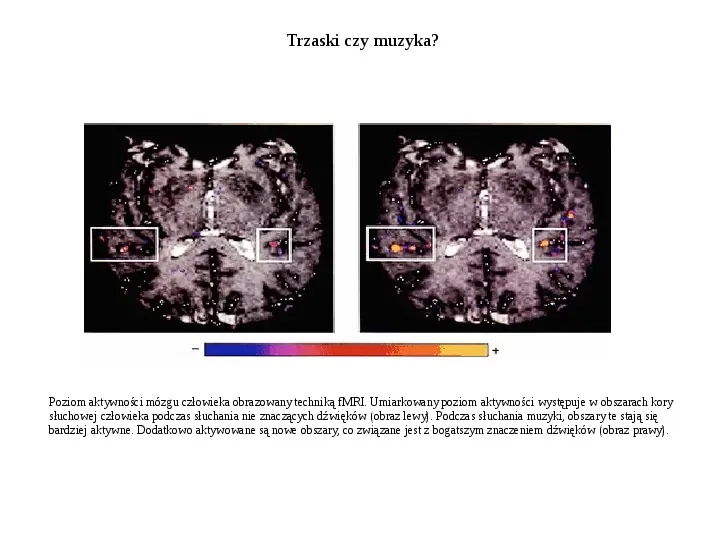
Trzaski czy muzyka? Poziom aktywności mózgu człowieka obrazowany techniką fMRI. Umiarkowany poziom aktywności występuje w obszarach kory słuchowej człowieka podczas słuchania nie znaczących dźwięków (obraz lewy). Podczas słuchania muzyki, obszary te stają się bardziej aktywne. Dodatkowo aktywowane są nowe obszary, co związane jest z bogatszym znaczeniem dźwięków (obraz prawy).
Dane:
- Liczba slajdów: 51
- Rozmiar: 4.95 MB
- Ilość pobrań: 59
- Ilość wyświetleń: 7030




