Zobacz slidy



























Treść prezentacji
Slide 1
METABOLIZM LIPIDÓW Iwonna Rahden-Staroń Katedra i Zakład Biochemii
Slide 2
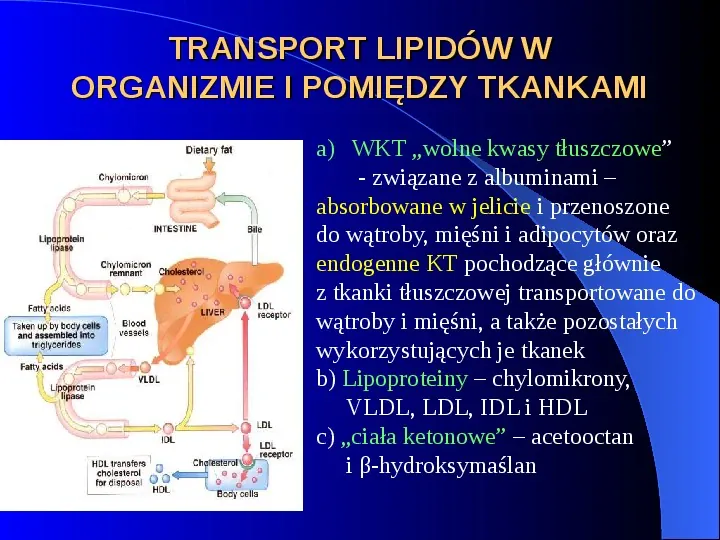
TRANSPORT LIPIDÓW W ORGANIZMIE I POMIĘDZY TKANKAMI a) WKT wolne kwasy tłuszczowe - związane z albuminami absorbowane w jelicie i przenoszone do wątroby, mięśni i adipocytów oraz endogenne KT pochodzące głównie z tkanki tłuszczowej transportowane do wątroby i mięśni, a także pozostałych wykorzystujących je tkanek b) Lipoproteiny chylomikrony, VLDL, LDL, IDL i HDL c) ciała ketonowe acetooctan i β-hydroksymaślan
Slide 3
Chylomikrony Transportują lipidy diety z enterocytów do tkanek organizmu Większość lipidów to triglicerydy Mało białka ApoA-I, ApoA-II, ApoB-48, ApoC Resynteza: TAG, EstryCh i Fosfolipidy Apolipoproteina B-48 Wit. rozp. tłuszczach do limfy do krwi
Slide 4
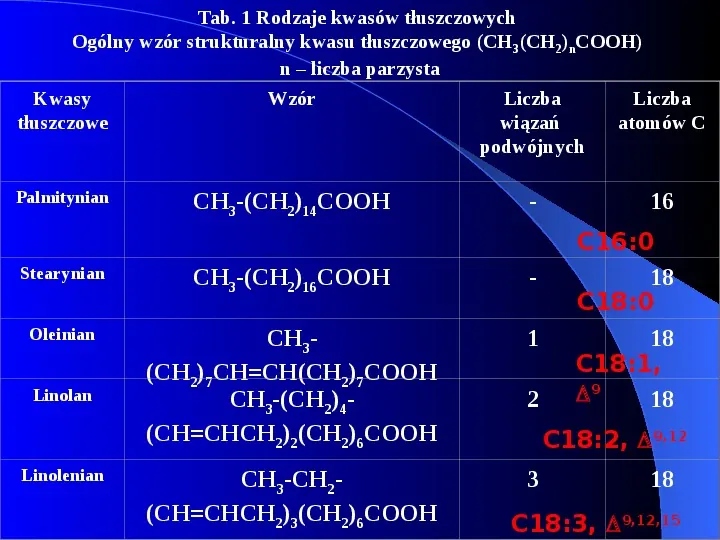
Tab. 1 Rodzaje kwasów tłuszczowych Ogólny wzór strukturalny kwasu tłuszczowego (CH 3(CH2)nCOOH) n liczba parzysta Kwasy tłuszczowe Wzór Liczba wiązań podwójnych Liczba atomów C Palmitynian CH3-(CH2)14COOH - 16 Stearynian CH3-(CH2)16COOH - Oleinian CH3(CH2)7CHCH(CH2)7COOH CH3-(CH2)4(CHCHCH2)2(CH2)6COOH 1 CH3-CH2(CHCHCH2)3(CH2)6COOH 3 Linolan Linolenian 2 C16:0 18 C18:0 18 C18:1, 9 18 C18:2, 9,12 18 C18:3, 9,12,15
Slide 5
Kwasy Tłuszczowe jako źródło energii α- oksydacja retikulum endoplazmatyczne, mitochondria służy prawdopodobnie utlenianiu nietypowych KT (np. z gr. CH3) [utl. C 2 poprzez OH i usunięcia C 1 jako CO2, z utlenieniem OH do COOH skrócenie o 1C β - oksydacja mitochondria energia, udział karnityny, acetylo-CoA; peroksysomy skracanie KT n 20C, rozgałęzionych, hydroksylowanych, brak udziału karnityny i zysku energii ω oksydacja retikulum endoplazmatyczne metabolizm nietypowych KT (w tym hydroksykwasów poprzez utlenienie ostatniego węgla (CH3- ) do grupy COOH, powstawanie krótkich kwasów dikarboksylowych
Slide 6
Synteza kwasów tłuszczowych -lipogeneza acetylo-CoA 7 malonylo-CoA 14 (NADPH H ) Syntaza KT kwas palmitynowy (16 C) 7 CO2 14 NADP 8 CoA 6 H2O AcetyloCoA łączy się z malonyloCoA 7 kolejnych etapów katalizowanych przez wielofunkcyjny enzym syntazę kwasów tłuszczowych Proces aktywny w tkance tłuszczowej, w gruczołach mlekowych w czasie laktacji Granica - 16 C (palmitynian) Kluczowymi enzymami regulacyjnym są: karboksylaza acetyloCoA (powstaje malonyloCoA) syntaza kwasów tłuszczowych
Slide 7
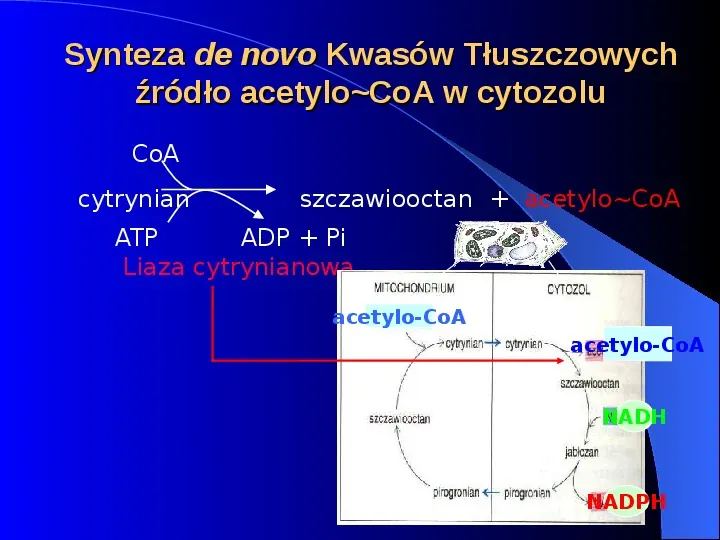
Synteza de novo Kwasów Tłuszczowych źródło acetyloCoA w cytozolu CoA cytrynian szczawiooctan acetyloCoA ADP Pi ATP Liaza cytrynianowa acetylo-CoA acetylo-CoA NADH NADPH
Slide 8
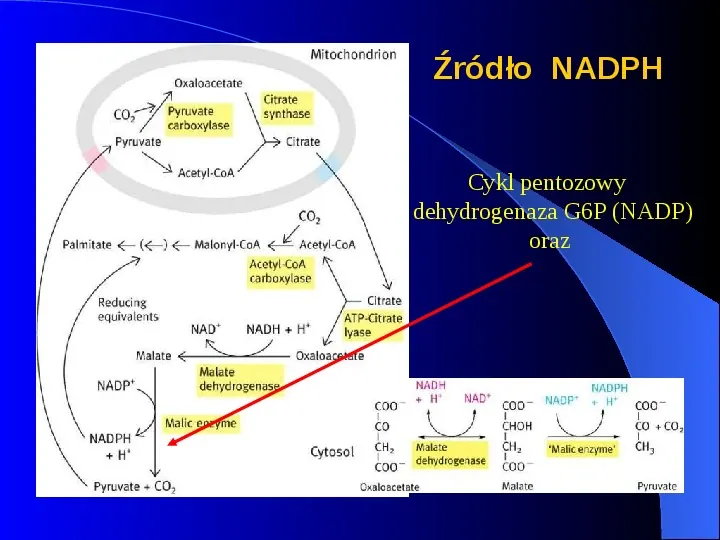
Źródło NADPH Cykl pentozowy dehydrogenaza G6P (NADP) oraz
Slide 9
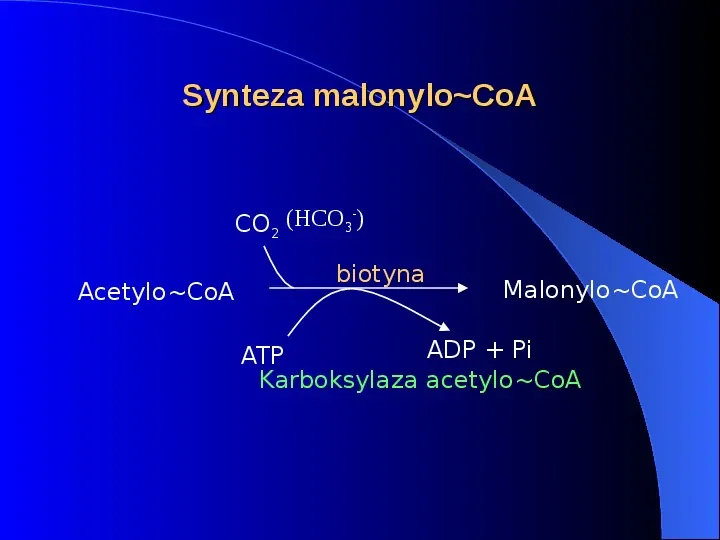
Synteza malonyloCoA CO2 (HCO3 ) - AcetyloCoA biotyna MalonyloCoA ADP Pi ATP Karboksylaza acetyloCoA
Slide 10
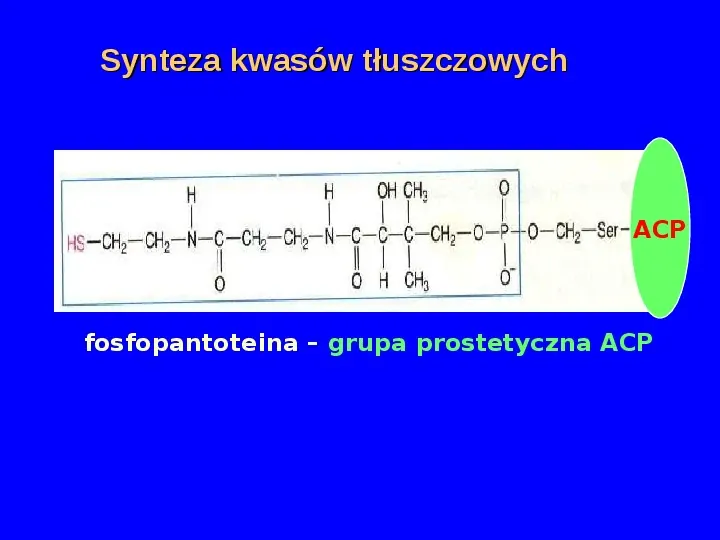
Synteza kwasów tłuszczowych ACP fosfopantoteina grupa prostetyczna ACP
Slide 11
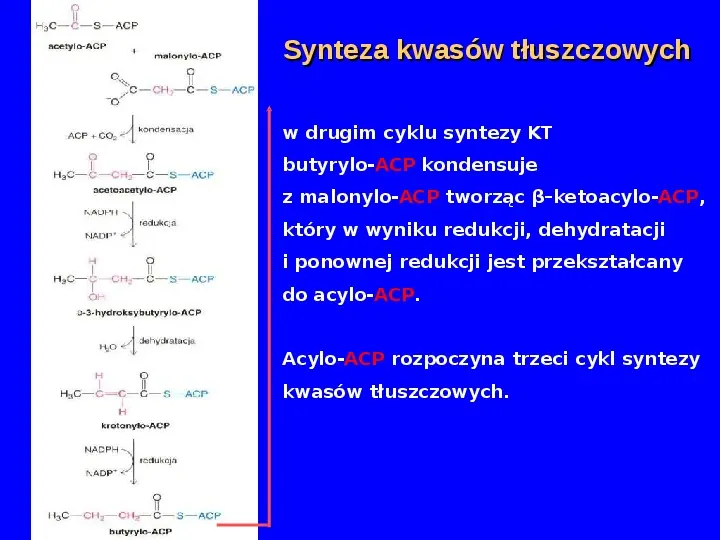
Synteza kwasów tłuszczowych w drugim cyklu syntezy KT butyrylo-ACP kondensuje z malonylo-ACP tworząc βketoacylo-ACP, który w wyniku redukcji, dehydratacji i ponownej redukcji jest przekształcany do acylo-ACP. Acylo-ACP rozpoczyna trzeci cykl syntezy kwasów tłuszczowych.
Slide 12
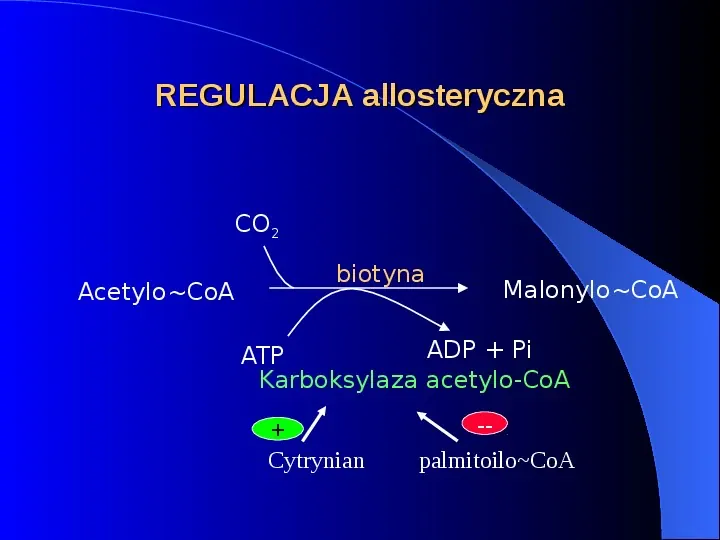
REGULACJA allosteryczna CO2 AcetyloCoA biotyna MalonyloCoA ADP Pi ATP Karboksylaza acetylo-CoA Cytrynian -palmitoiloCoA
Slide 13
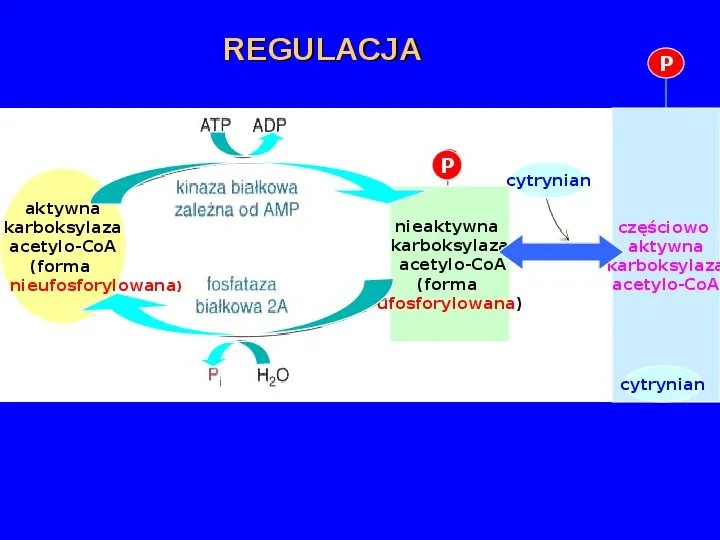
REGULACJA P P aktywna karboksylaza acetylo-CoA (forma nieufosforylowana) cytrynian nieaktywna karboksylaza acetylo-CoA (forma ufosforylowana) częściowo aktywna karboksylaza acetylo-CoA cytrynian
Slide 14
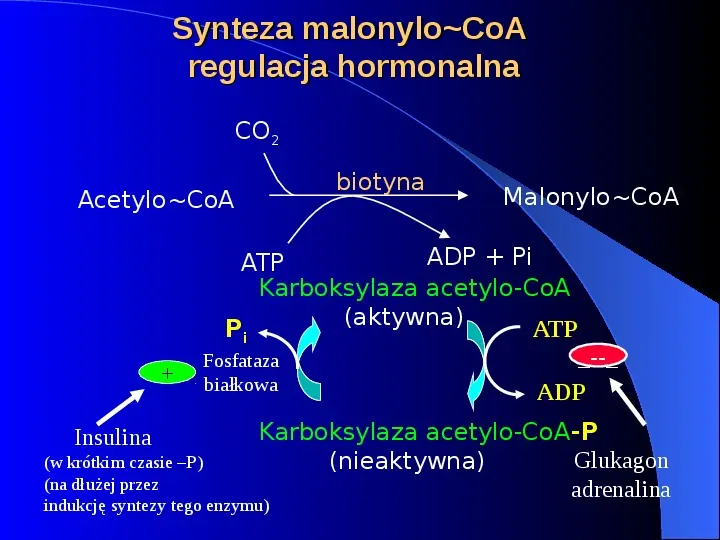
Synteza malonyloCoA regulacja hormonalna CO2 biotyna AcetyloCoA Insulina MalonyloCoA ADP Pi ATP Karboksylaza acetylo-CoA (aktywna) Pi ATP -- Fosfataza białkowa ADP Karboksylaza acetylo-CoA-P Glukagon (nieaktywna) adrenalina (w krótkim czasie P) (na dłużej przez indukcję syntezy tego enzymu)
Slide 15
Wydłużanie kwasów tłuszczowych i wprowadzanie wiązań podwójnych U Eukariontów wydłużanie KT dłuższych niż palmitynian (C16) i tworzenie wiązania podwójnego są katalizowane przez enzymy zlokalizowane na powierzchni gładkiego retikulum endoplazmatycznego. Donorem jednostek dwuwęglowych jest malonylo-CoA, a wydłużny KT jest związany z CoA, a nie z ACP. Wprowadzenie wiązania podwójnego wymaga aktywności 3 enzymów: reduktazy cytochromu b5 [NADH, O2], cytochromu b5 i desaturazy. Δ9, Δ6, Δ5, Δ4
Slide 16
Wiązania podwójne W organizmach ssaków nie występują enzymy zdolne do tworzenia wiązań podwójnych w położeniu dalszym niż przy węglu C9. Dlatego ssaki nie potrafią syntetyzować NNKT kwas linolowy C18: cis - Δ9, Δ12 (ω-6) do syntezy kwasu arachidonowego kwas linolenowy C18: cis - Δ9, Δ12, Δ15 (ω-3) przekształcany do kwasu dokozaheksaenowego (DHA) C 22:6 kwasu eikozapentaenowego (EPA) C 20:5
Slide 17
Funkcje Niezbędnych Kwasów Tłuszczowych Eikozanoidy Lipidowe mediatory zapalenia syntetyzowane z kwasu arachidonowego i EPA Zawierają Prostaglandyny Prostacykliny Tromboksany Leukotrieny
Slide 18
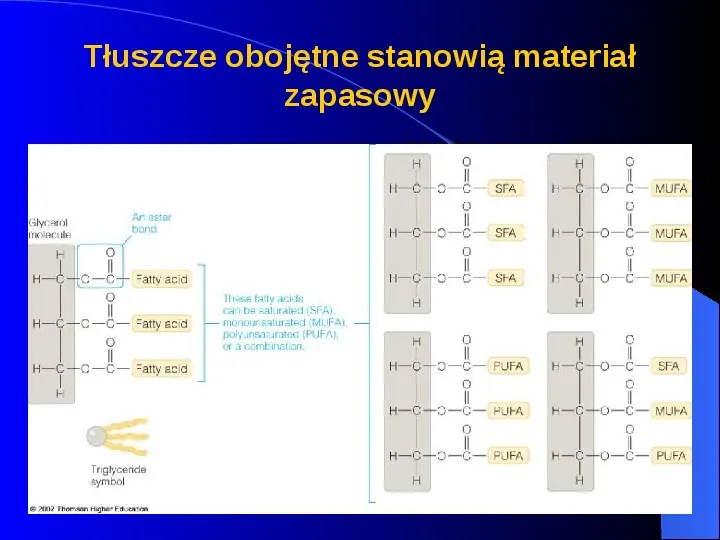
Tłuszcze obojętne stanowią materiał zapasowy
Slide 19
Funkcje Triglicerydów Dostarczają niezbędne KT do funkcjonowania organizmu Produkcja energii Izolacja Ochrona
Slide 20
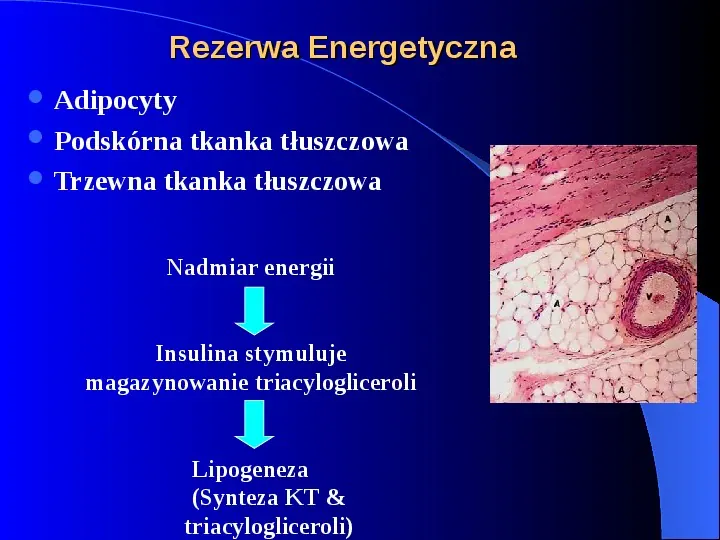
Rezerwa Energetyczna Adipocyty Podskórna tkanka tłuszczowa Trzewna tkanka tłuszczowa Nadmiar energii Insulina stymuluje magazynowanie triacylogliceroli Lipogeneza (Synteza KT triacylogliceroli)
Slide 21
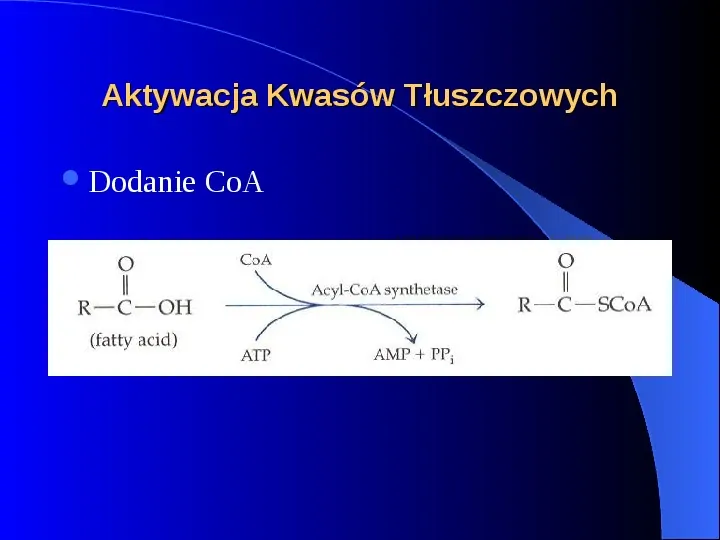
Aktywacja Kwasów Tłuszczowych Dodanie CoA
Slide 22
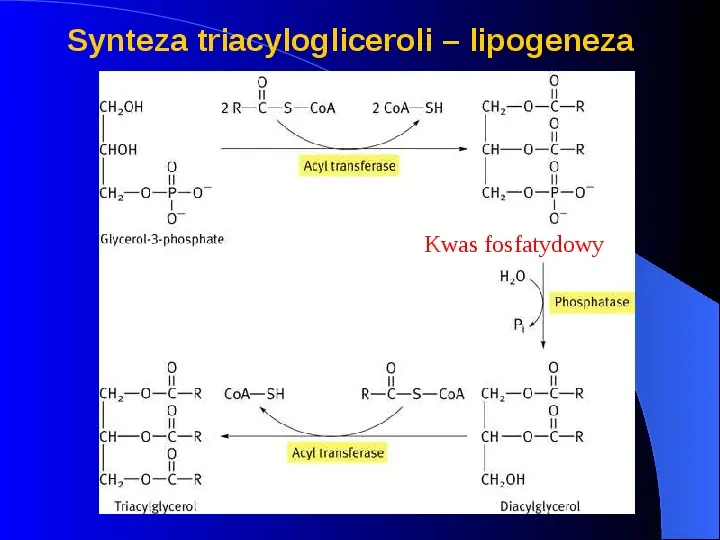
Synteza triacylogliceroli lipogeneza Kwas fosfatydowy
Slide 23
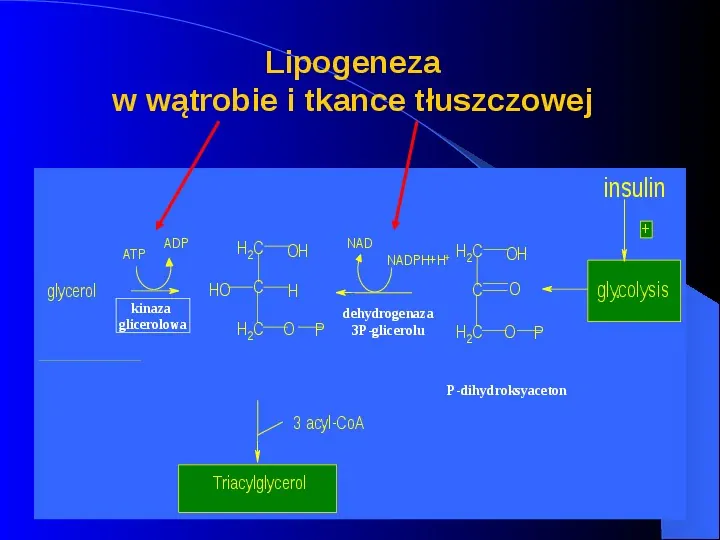
Lipogeneza w wątrobie i tkance tłuszczowej insulin ATP glycerol ADP glycerol kinaza kinase glicerolowa minor pathway H2C HO C H2C NAD OH H O P NADPHH glycerol-3P dehydrogenaza dehydrogenase 3P-glicerolu sn-glycerol -3P 3 acyl-CoA Triacylglycerol H2C OH C O H2C O glycolysis P dihydroxyacetonephosphate P-dihydroksyaceton major pathway
Slide 24
Metabolizm Lipidów Złożonych
Slide 25
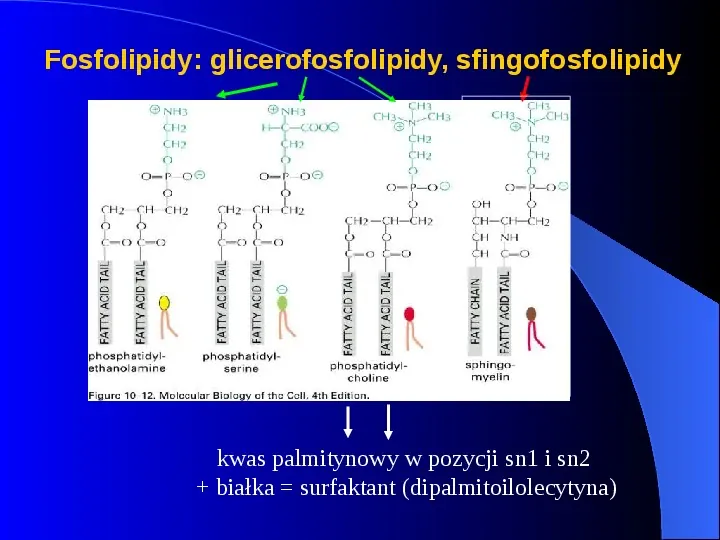
Fosfolipidy: glicerofosfolipidy, sfingofosfolipidy kwas palmitynowy w pozycji sn1 i sn2 białka surfaktant (dipalmitoilolecytyna)
Slide 26
Synteza Lipidów Złożonych Glicerofosfolipidy [1] kwas fosfatydowy CTP CDP-diacyloglicerol PPi CDP-diacyloglicerol cholina (etanoloamina, seryna, inozytol) lecytyna (fosfatydyloetanoloamina, fosfatydyloseryna, 4,5-difosfoinozytyd) CMP [2] DAG CDP-cholina (CDP-etanoloamina, CDP-seryna, CDP-inozytol) lecytyna (fosfatydyloetanoloamina, fosfatydyloseryna, 4,5-difosfoinozytyd) CMP [3] wzajemne przekształcenia fosfolipidów pomiędzy sobą udział m.in. metylacji i SAM jako dawcy grup CH3 fosfatydyloetanoloamina 3 SAM fosfatydylocholina (lecytyna)
Slide 27
Synteza Lipidów Złożonych Sfingofosfolipidy palmityloCoA seryna sfingozyna sfingozyna acyloCoA ceramid Ceramid CDP-cholina sfingomielina Ceramid UDP-glukoza lub UDP-galaktoza cerebrozydy Cerebrozydy UDP-monosacharyd (CMP-NANA) gangliozydy Gangliozydy oparte na sfingozynie to co odróżnia grupy A, B i 0 krwi człowieka
Slide 28
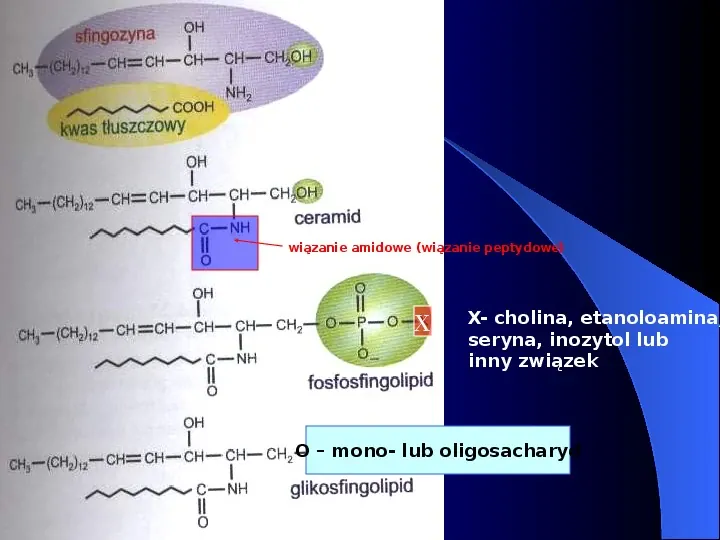
wiązanie amidowe (wiązanie peptydowe) X X- cholina, etanoloamina, seryna, inozytol lub inny związek O mono- lub oligosacharyd
Slide 29
Sterole Estry Steroli Steroid cyklopentanoperhydrofenantren
Slide 30
Synteza Cholesterolu Całkowita dzienna synteza cholesterol ok. 700mg24 h Zaleca się spożycie 300 mgdzień W wątrobie powstaje 10-20 endogennego cholesterolu, 10 w jelitach. Wszystkie komórki jądrzaste są zdolne do syntezy cholesterolu. Enzymy syntezy znajdują się we frakcji mikrosomalnej i cytosolowej komórki
Slide 31
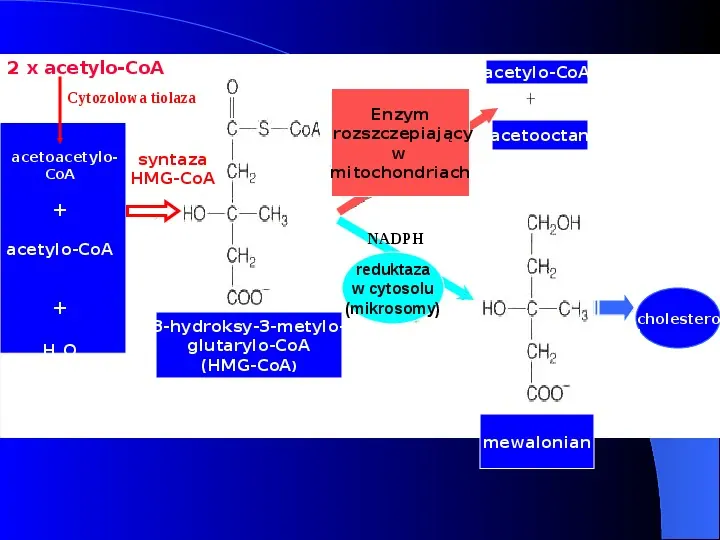
2 x acetylo-CoA Cytozolowa tiolaza acetoacetylo- syntaza CoA HMG-CoA acetylo-CoA Enzym rozszczepiający w mitochondriach acetooctan acetylo-CoA H2O NADPH reduktaza w cytosolu (mikrosomy) 3-hydroksy-3-metyloglutarylo-CoA (HMG-CoA) cholestero l mewalonian
Slide 32
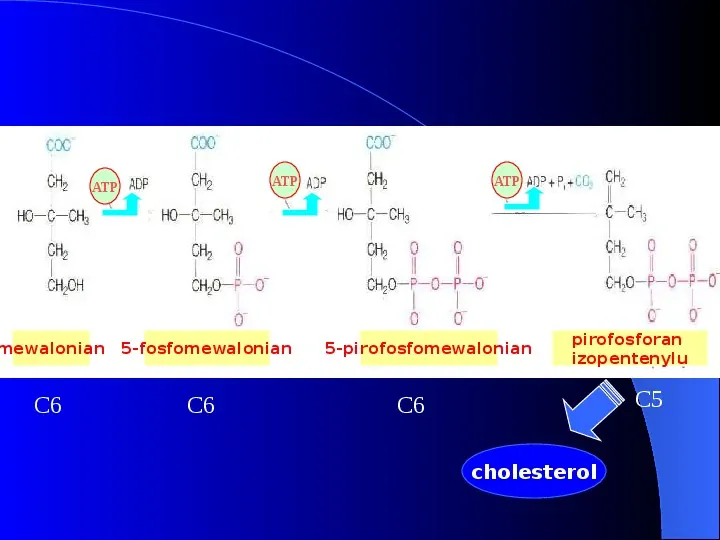
ATP ATP mewalonian C6 5-fosfomewalonian C6 ATP 5-pirofosfomewalonian pirofosforan izopentenylu C5 C6 cholesterol
Slide 33
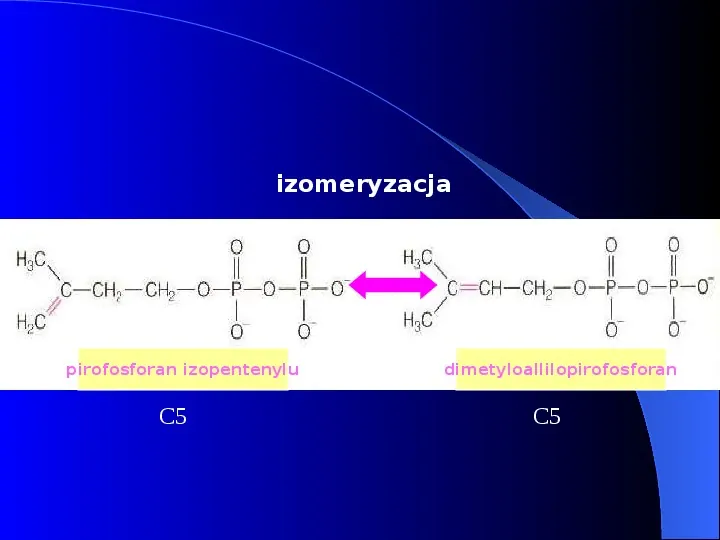
izomeryzacja pirofosforan izopentenylu C5 dimetyloallilopirofosforan C5
Slide 34
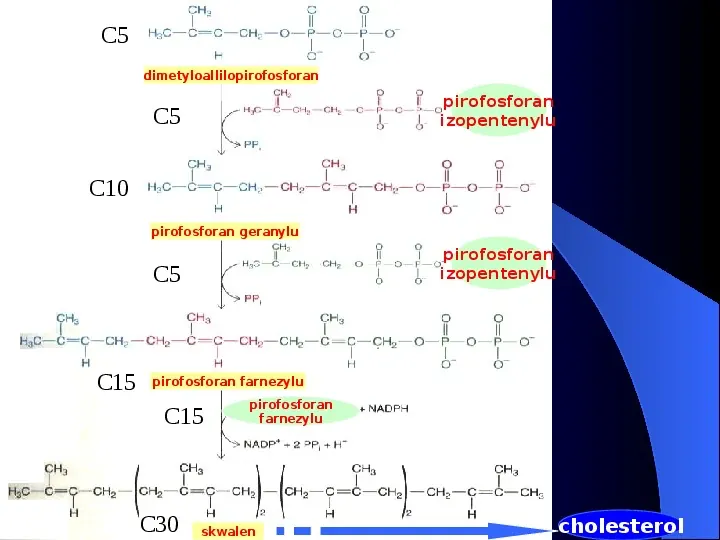
C5 dimetyloallilopirofosforan pirofosforan izopentenylu C5 C10 pirofosforan geranylu pirofosforan izopentenylu C5 C15 pirofosforan farnezylu C15 C30 pirofosforan farnezylu skwalen cholesterol
Slide 35
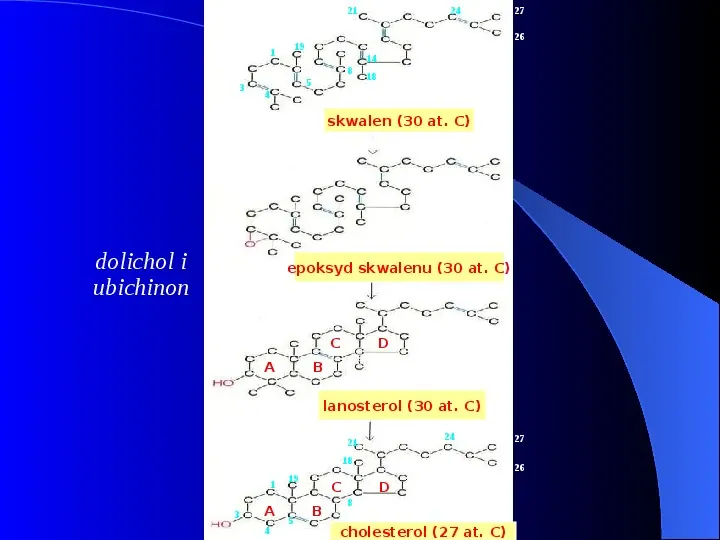
21 27 26 19 1 14 8 3 24 5 18 4 skwalen (30 at. C) dolichol i ubichinon epoksyd skwalenu (30 at. C) C D B A lanosterol (30 at. C) 3CH3 24 21 18 1 3 A 4 19 5 C B 27 26 D 8 cholesterol (27 at. C)
Slide 36
Regulacja syntezy cholesterolu Kluczowy enzym - reduktaza HMG-CoA. [1] hamowanie allosteryczne przez metaboliczne pochodne mewalonianu i cholesterolu [2] hamowanie przez modyfikacje kowalencyjna fosforylacja zależna od glukagonu (kaskada z udziałem cAMP i kinazy białkowej A) P-enzym nieaktywny [3] aktywacja przez uzależnioną od insuliny defosforylację (aktywny) (fosfataza fosfobiałkowa); insulina stymuluje ekspresję genu reduktazy Statyny Obniżają poziom cholesterolu we krwi przez hamowanie syntezy cholesterolu (kompetycyjny inhibitor reduktazy HMG-CoA).
Slide 37
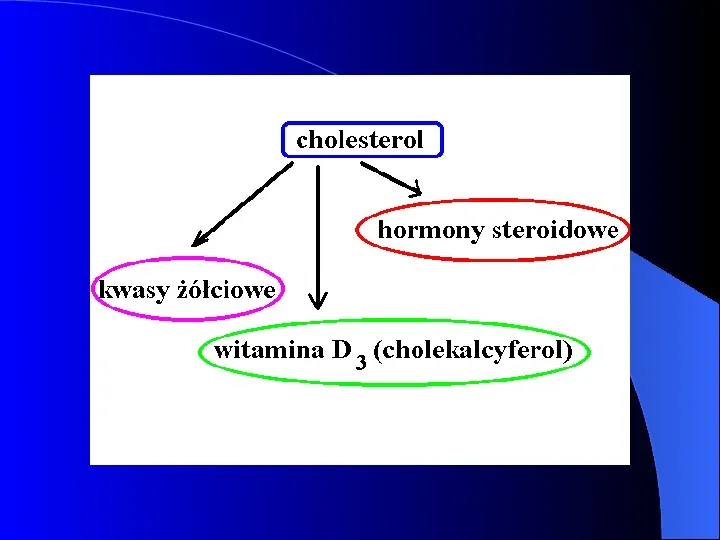
Wykorzystanie cholesterolu w organizmie - główny składnik błony cytoplazmatycznej - nadmiar - miażdżyca - istotny problem usuwanie synteza kwasów żółciowych główna droga (0.5 gdobę) synteza hormonów steroidowych - kora nadnerczy, gonady synteza witaminy D3 wątroba, nerka Estry cholesterolu forma magazynowania cholesterolu w komórce, zasadnicza postać przenoszenia cholesterolu w lipoproteinach.
Slide 38
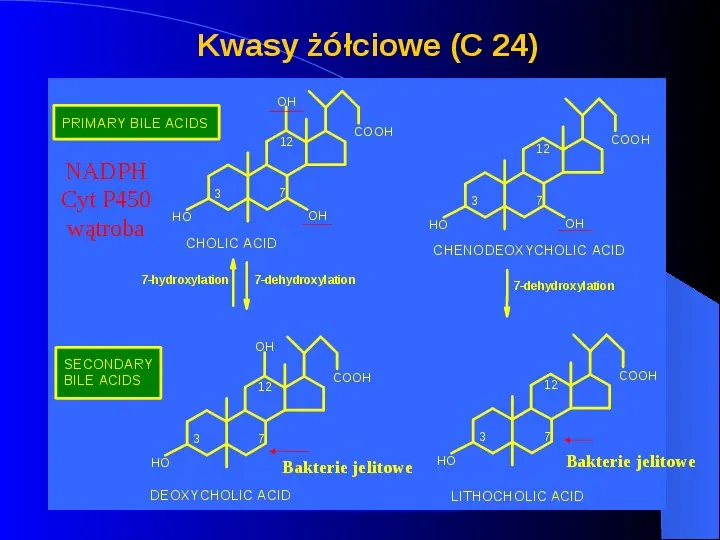
Cholesterol Cholesterol NIE jest związkiem dostarczającym energii Jest transportowany do wątroby i usuwany w żółci 55 jako cholesterol 45 jako kwasy żółciowe Kwas cholowy i chenodeoksycholowy Skonjugowane z glicyną lub tauryną Glikocholowy lub taurocholowy kwas
Slide 40
Kwasy żółciowe (C 24) OH PRIMARY BILE ACIDS COOH 12 NADPH Cyt P450 wątroba 7 3 OH HO CHOLIC ACID 7-hydroxylation 7 3 OH HO COOH 12 CHENODEOXYCHOLIC ACID 7-dehydroxylation 7-dehydroxylation OH SECONDARY BILE ACIDS 3 HO COOH 12 3 7 Bakterie jelitowe DEOXYCHOLIC ACID COOH 12 HO 7 Bakterie jelitowe LITHOCHOLIC ACID
Slide 41
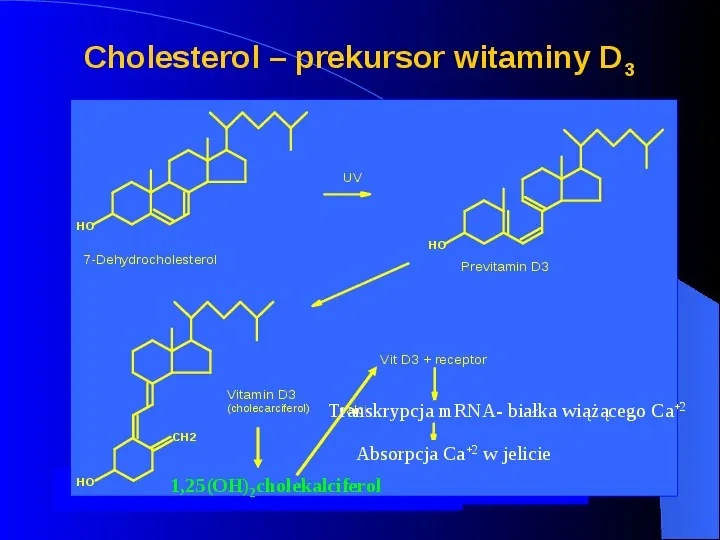
Cholesterol prekursor witaminy D3 UV HO HO 7-Dehydrocholesterol Previtamin D3 Vit D3 receptor Vitamin D3 (cholecarciferol) CH2 HO calcium binding mRNAprotein mRNA transcription Transkrypcja białka wiążącego Ca2 Absorpcja Ca2inwthejelicie absorption of calcium intestine 1,25(OH)2cholekalciferol
Slide 42
Hormony steroidowe Cholesterol (C27) ACTH NADPH Cyt. P450, O2 pregnenolon (C21) progestageny (C21) (progesteron - prekursor) glikokortykosteroidy (C21) androgeny (C19) kortyzol mineralokortykosteroidy (C21) aldosteron estrogeny (C18) ACTH aktywacja cyklazy adenylanowej (cAMP ), zwiększenie akt. PKA w tkance rdzenia akt. enzymu odcinającego łańcuch boczny, który przekształca cholesterol do pregnenolonu. Zasadniczy efekt fizjologiczny ACTH to produkcja glikokortykosteroidów.
Slide 43
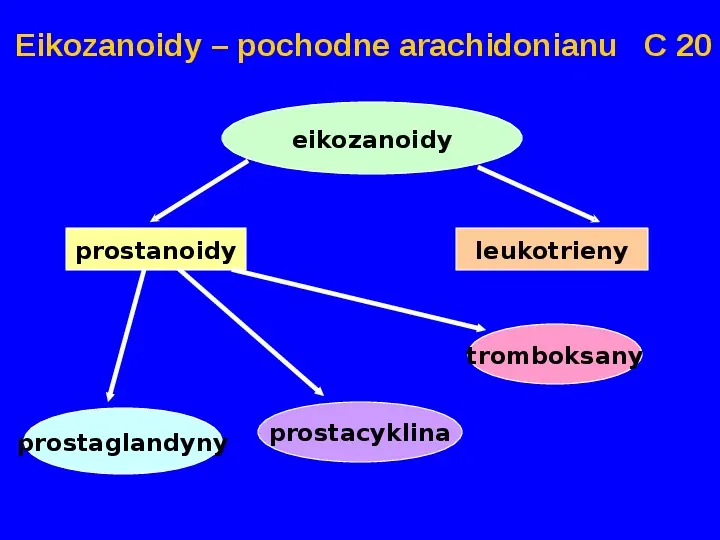
Eikozanoidy pochodne arachidonianu C 20 eikozanoidy prostanoidy leukotrieny tromboksany prostaglandyny prostacyklina
Slide 44
Degradacja fosfolipidów a. fosfolipazy A1 Sok trzustkowy, tkanki Uwalnia kw. arachidonowy. Obecna w jadzie węży i O pszczół R -C 2 A2 tkanki O CH2 - O - C R1 O -CH CH2 O - P - X Lizosomy wątroby -toksyna Clostridium i in. Bakterii. Uwalnia wtórne przekaźniki. C D głównie rośliny
Slide 45
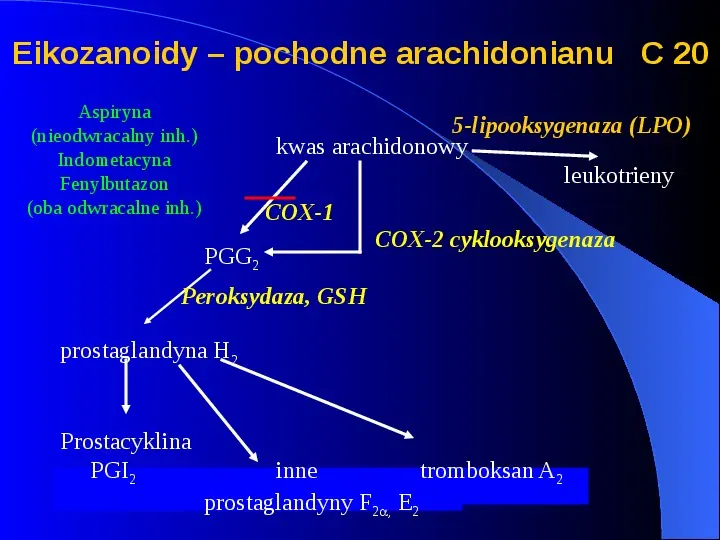
Eikozanoidy pochodne arachidonianu C 20 Aspiryna (nieodwracalny inh.) Indometacyna Fenylbutazon (oba odwracalne inh.) 5-lipooksygenaza (LPO) kwas arachidonowy leukotrieny COX-1 PGG2 COX-2 cyklooksygenaza Peroksydaza, GSH prostaglandyna H2 Prostacyklina PGI2 inne tromboksan A2 prostaglandyny F2, E2
Slide 46
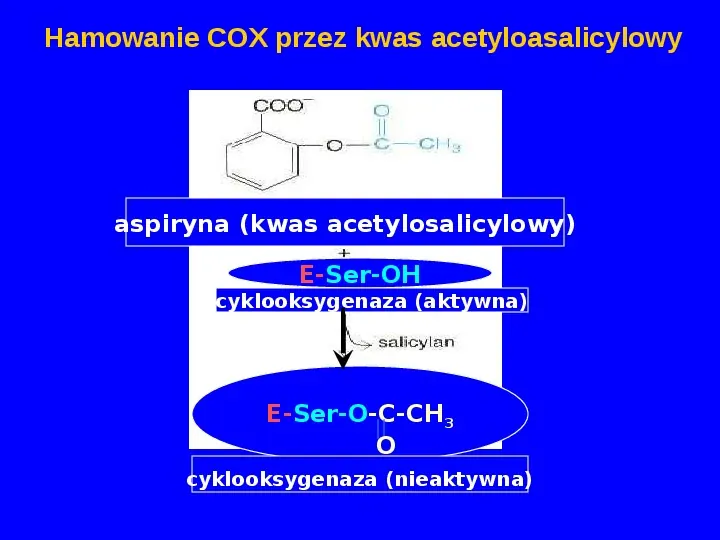
Hamowanie COX przez kwas acetyloasalicylowy aspiryna (kwas acetylosalicylowy) E-Ser-OH cyklooksygenaza (aktywna) E-Ser-O-C-CH3 O cyklooksygenaza (nieaktywna)
Slide 47
Katabolizm Lipidów mobilizacja zapasów tłuszczu i utlenianie KT Lipoliza Transport KT Pobieranie KT, aktywacja i utlenianie
Slide 48
Lipoliza Rozpad triacylogliceroli do glicerolu 3 kwasów tłuszczowych (wolnych) Lipaza hormono-wrażliwa w tkance tłuszczowej Lipaza lipoproteinowa w naczyniach krwionośnych Wolne kwasy tłuszczowe zwane też niezestryfikowanymi kwasami tłuszczowymi
Slide 49
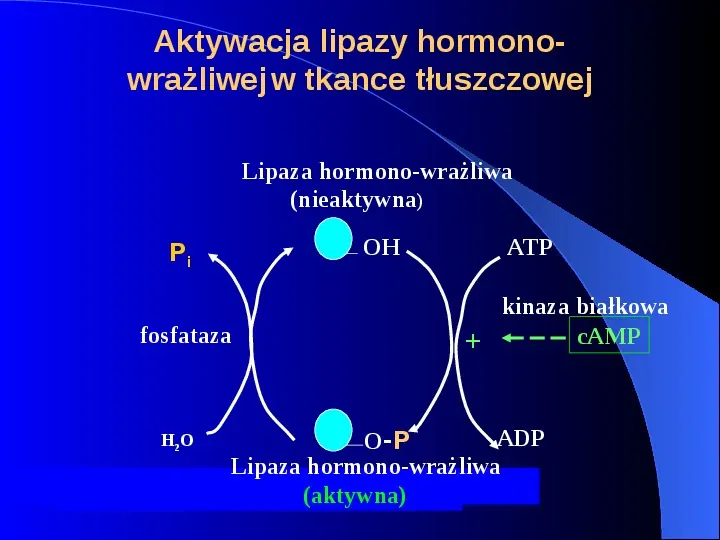
Aktywacja lipazy hormonowrażliwej w tkance tłuszczowej Lipaza hormono-wrażliwa (nieaktywna) OH Pi fosfataza H2 O ATP kinaza białkowa cAMP ADP O-P Lipaza hormono-wrażliwa (aktywna)
Slide 50
Źródło Energii Lipoliza Rozpad triglicerydów do KT glicerolu Stymulowana przez: Niski poziom insuliny podczas hypoglikemii Ćwiczenia sportowe Stres fizjologiczny Lipaza hormono-wrażliwa Enzym, który katalizuje hydrolizę 1-go wiązania estrowego w TAG Mobilizuje KT z tkanki tłuszczowej
Slide 51
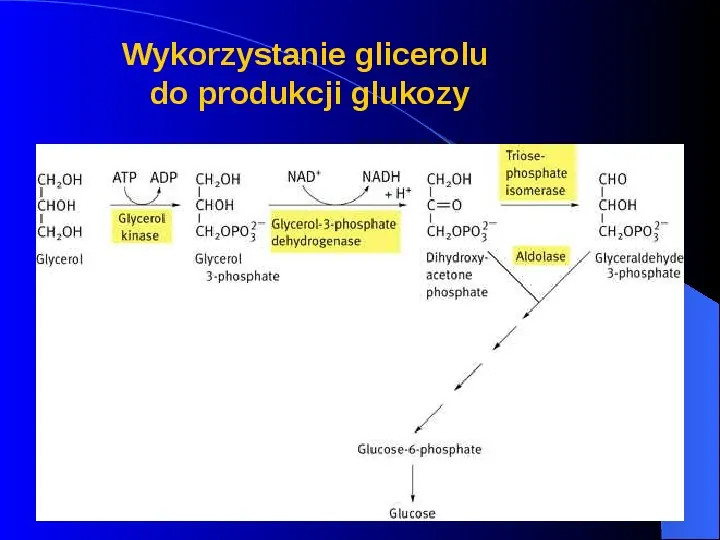
Wykorzystanie glicerolu do produkcji glukozy
Slide 52
Degradacja sfingomieliny b) Sfingomielinaza Enzym lizosomalny Uwalnia P-cholinę Zostaje ceramid ----- ceramidaza Odpowiedź na stres Sfingozyna hamuje kinazę białkowa C Choroba Niemana-Picka sfingomielina nie jest rozkładana.
Slide 53
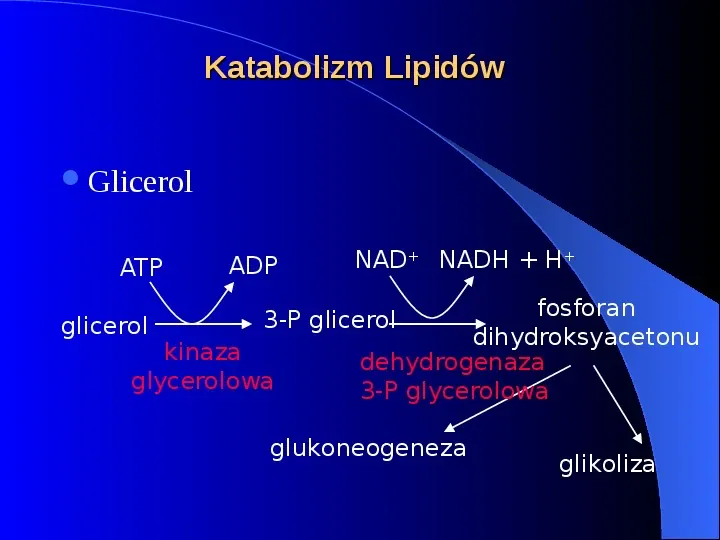
Katabolizm Lipidów Glicerol ATP glicerol ADP NAD NADH H fosforan dihydroksyacetonu dehydrogenaza 3-P glycerolowa 3-P glicerol kinaza glycerolowa glukoneogeneza glikoliza
Slide 54
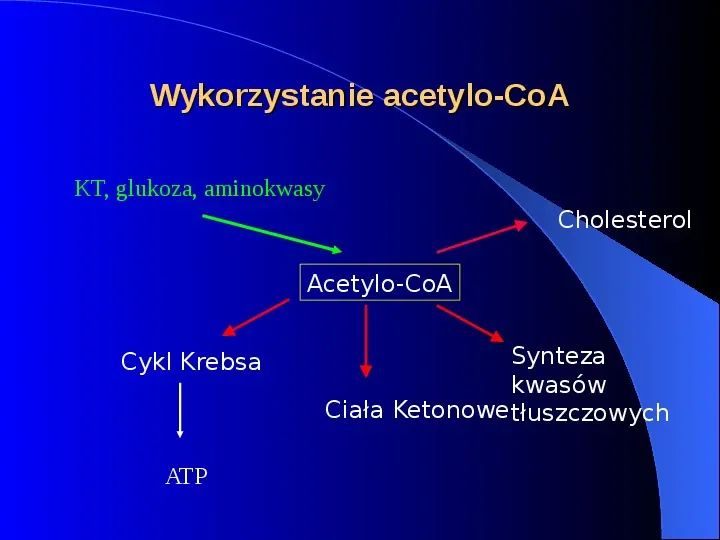
Wykorzystanie acetylo-CoA KT, glukoza, aminokwasy Cholesterol Acetylo-CoA Cykl Krebsa ATP Synteza kwasów Ciała Ketonowe tłuszczowych
Slide 55
Źródło Energii 1 g KT dostarcza 9 kcal Ketogeneza Występuje, gdy jest ograniczony dostęp glukozy do organizmu Powstałe ciała ketonowe Są wykorzystane przez mózg, serce, mięśnie szkieletowe, nerki Oszczędzają białka
Slide 56
Ketoza Występuje, gdy synteza ciał ketonowych przewyższa wykorzystanie Choroba - ketoacidoza Cukrzyca typu 1 (zależna od insuliny) Insulina Glukagon Lipoliza Wolne kwasy tłuszczowe w osoczu Wyrzut ciał ketonowych z wątroby Ketoacidoza ketonemiaketonuria
Slide 57
Ketony β-oksydacja prowadzi do produkcji znacznych ilości acetylo-CoA Jeśli brakuje szczawiooctanu, wtedy acetylo-CoA nie przechodzi przez cykl Krebsa i jest przekształcany do ciał ketonowych Niski poziom glikolizy prowadzi do zmniejszonej produkcji szczawiooctanu
Slide 58
Wykorzystanie ciał ketonowych przez tkanki obwodowe Acetoocatan i β-hydroksymaślan nie są dalej utleniane w wątrobie lecz wędrują do tkanek obwodowych, gdzie mogą być przekształcone z powrotem do acetylo-CoA i utlenione via cykl Krebsa. mózg, mięśnie, nerki NAD 3-hydroksymaślan bursztynylo-CoA bursztynian X dh. 3-hydroksymaślanu NADHH X acetooctan tioforaza acetoacetylo-CoA 2 acetylo-CoA
Slide 59
Ciała ketonowe alternatywne źródło energii dla komórek Acetooctan, β-hydroksymaślan, aceton Acetylo-CoA acetylo-CoA acetoacetylo-CoA Acetoacetylo-CoA CoA acetooctan Acetooctan może być przekształcony do acetonu i -hydroksymaślanu β
Dane:
- Liczba slajdów: 59
- Rozmiar: 3.33 MB
- Ilość pobrań: 306
- Ilość wyświetleń: 9358